胃の腺の上皮。 胃の細胞。 胃ホルモン。 胃自身の腺の細胞小腸での消化
調子キュー腸は、条件付きで十二指腸、空腸、回腸の3つのセクションに分けられます。 小腸の長さは6メートルで、主に植物性食品を摂取する人では12メートルに達することがあります。
小腸の壁はで構成されています 4シェル:粘液、粘膜下組織、筋肉および漿液。
小腸の粘膜には 自分の安堵、腸のひだ、腸絨毛、腸陰窩が含まれます。
腸のひだ粘膜と粘膜下組織によって形成され、本質的に円形です。 ケルクリング皺は十二指腸で最も高くなります。 小腸の過程で、ケルクリング皺の高さは減少します。
腸絨毛粘膜の指のような成長です。 十二指腸では、腸絨毛は短く幅が広く、小腸に沿って高く薄くなります。 腸のさまざまな部分の絨毛の高さは0.2〜1.5mmに達します。 絨毛の間で3-4腸陰窩を開きます。
腸陰窩小腸のコースに沿って増加する粘膜のそれ自身の層への上皮のくぼみです。
小腸の最も特徴的な形成は、腸絨毛と腸陰窩であり、これらは表面を大幅に増加させます。
表面から、小腸の粘膜(絨毛と陰窩の表面を含む)は単層の角柱状の上皮で覆われています。 腸上皮の寿命は24時間から72時間です。 固形食は、シャロンを生成する細胞の死を加速し、陰窩上皮細胞の増殖活性の増加につながります。 現代のアイデアによると、 生成ゾーン腸上皮の1つは陰窩の底であり、すべての上皮細胞の12〜14%が合成期にあります。 生命活動の過程で、上皮細胞は陰窩の深さから絨毛の上部に徐々に移動し、同時に多くの機能を実行します:増殖し、腸で消化された物質を吸収し、粘液と酵素を腸の内腔に分泌します。 腸内の酵素の分離は、主に腺細胞の死とともに起こります。 絨毛の上部に上昇する細胞は拒絶され、腸の内腔で崩壊し、そこで酵素を消化器系の粥状液に与えます。
腸細胞の中には、自分のプレートからここに浸透し、Tリンパ球(細胞毒性、Tメモリー細胞、ナチュラルキラー)に属する上皮内リンパ球が常にあります。 上皮内リンパ球の含有量は、さまざまな病気や免疫障害で増加します。 腸上皮いくつかのタイプの細胞要素(腸細胞)が含まれます:境界、ゴブレット、境界なし、房状、内分泌、M細胞、パネート細胞。
境界細胞(円柱状)腸上皮細胞の主要な集団を構成します。 これらの細胞は角柱状であり、頂端面にはゆっくりと収縮する能力を持つ多数の微絨毛があります。 事実、微絨毛には細いフィラメントと微小管が含まれています。 各微絨毛には、中央にアクチンマイクロフィラメントの束があり、片側で絨毛の頂点のプラスモレンマに接続されており、基部では、末端ネットワーク(水平方向のマイクロフィラメント)に接続されています。 この複合体は、吸収中に微絨毛の収縮を確実にします。 絨毛の境界細胞の表面には800から1800の微絨毛があり、陰窩の境界細胞の表面には225の微絨毛しかありません。 これらの微絨毛は縞模様の境界を形成します。 表面から、微絨毛は糖衣の厚い層で覆われています。 境界細胞の場合、細胞小器官の極性配列が特徴的です。 核は基底部にあり、その上にゴルジ装置があります。 ミトコンドリアも頂端極に局在しています。 それらはよく発達した顆粒状および無顆粒状の小胞体を持っています。 細胞の間に、細胞間空間を閉じる終板があります。 細胞の頂端部には、細胞表面に平行なフィラメントのネットワークからなる明確な末端層があります。 ターミナルネットワークは、アクチンとミオシンのマイクロフィラメントを含み、腸細胞の頂端部の側面の細胞間接触に接続されています。 末端ネットワークにマイクロフィラメントが関与することで、腸細胞間の細胞間ギャップが閉じられ、消化中にさまざまな物質が腸細胞に侵入するのを防ぎます。 微絨毛の存在は細胞表面を40倍に増加させ、それにより小腸の総表面が増加し、500mに達します。 微絨毛の表面には、胃液や腸液の酵素(ホスファターゼ、ヌクレオシドジホスファターゼ、アミノペプチダーゼなど)によって破壊されない分子の加水分解による切断を提供する多数の酵素があります。 このメカニズムは、膜または壁側消化と呼ばれます。
膜消化小分子の分裂のための非常に効果的なメカニズムだけでなく、加水分解と輸送のプロセスを組み合わせた最も先進的なメカニズムもあります。 微絨毛の膜に位置する酵素には二重の起源があります。それらは部分的に粥状液から吸着され、部分的に境界細胞の顆粒状小胞体で合成されます。 膜消化中に、ペプチドとグルコシド結合の80〜90%、トリグリセリドの55〜60%が切断されます。 微絨毛の存在は、腸の表面を一種の多孔質触媒に変えます。 微絨毛は収縮して弛緩することができ、それが膜消化のプロセスに影響を与えると考えられています。 グリコカリックスの存在と微絨毛の間の非常に小さなスペース(15〜20ミクロン)は、消化の無菌性を保証します。
切断後、加水分解生成物は微絨毛膜に浸透します。微絨毛膜は能動輸送と受動輸送の能力を持っています。
脂肪が吸収されると、最初に低分子量化合物に分解され、次に脂肪がゴルジ装置内および顆粒状小胞体の尿細管内で再合成されます。 この複合体全体が細胞の側面に輸送されます。 エキソサイトーシスにより、脂肪は細胞間空間に除去されます。
ポリペプチドおよび多糖鎖の切断は、微絨毛の原形質膜に局在する加水分解酵素の作用下で起こります。 アミノ酸と炭水化物は、能動輸送メカニズムを使用して、つまりエネルギーを使用して細胞に入ります。 その後、それらは細胞間空間に放出されます。
したがって、絨毛と陰窩に位置する境界細胞の主な機能は、腔内よりも数倍激しく進行する壁側消化であり、有機化合物の最終生成物への分解と加水分解生成物の吸収を伴います。
杯細胞大脳辺縁系腸細胞の間に単独で位置しています。 それらの含有量は、十二指腸から大腸への方向に増加します。 絨毛上皮よりも上皮に杯細胞陰窩が多い。 これらは典型的な粘液細胞です。 それらは、粘液の蓄積と分泌に関連する周期的な変化を示します。 粘液蓄積段階では、これらの細胞の核は細胞の基部に位置し、不規則または三角形の形状をしています。 細胞小器官(ゴルジ装置、ミトコンドリア)は核の近くにあり、よく発達しています。 同時に、細胞質は粘液の滴で満たされています。 分泌後、細胞のサイズが減少し、核が減少し、細胞質が粘液から解放されます。 これらの細胞は、粘膜の表面を湿らせるのに必要な粘液を生成します。これは、一方では粘膜を機械的損傷から保護し、他方では食物粒子の動きを促進します。 さらに、粘液は感染性の損傷から保護し、腸の細菌叢を調節します。
M細胞リンパ濾胞(グループと単一の両方)の局在領域の上皮に位置していますこれらの細胞は平らな形をしており、微絨毛の数が少ないです。 これらの細胞の頂端には多数のマイクロフォールドがあるため、「マイクロフォールドのある細胞」と呼ばれます。 マイクロフォールドの助けを借りて、それらは腸管腔から高分子を捕獲し、エンドサイトーシス小胞を形成することができ、それは原形質膜に輸送され、細胞間空間に放出され、次に粘膜固有層に放出される。 その後、リンパ球t。 抗原によって刺激されたpropriaはに移動します リンパ節それらが増殖して血流に入るところ。 末梢血を循環した後、固有層に再増殖し、そこでBリンパ球がIgA分泌形質細胞に変換されます。 したがって、腸腔から来る抗原はリンパ球を引き付け、それが腸のリンパ組織の免疫応答を刺激します。 M細胞では、細胞骨格の発達が非常に悪いため、上皮間リンパ球の影響下で容易に変形します。 これらの細胞はリソソームを持たないため、変化することなく小胞を介してさまざまな抗原を輸送します。 彼らは糖衣を欠いています。 ひだによって形成されたポケットにはリンパ球が含まれています。
房状の細胞それらの表面には、腸の内腔に突き出た長い微絨毛があります。 これらの細胞の細胞質には、滑らかな小胞体の多くのミトコンドリアと細管が含まれています。 それらの頂端部は非常に狭い。 これらの細胞は化学受容器として機能し、おそらく選択的吸収を行うと考えられています。
パネート細胞(好酸性の粒度を持つ外分泌細胞)は、グループまたは単独で陰窩の下部にあります。 それらの頂端部には、高密度の好酸性染色顆粒が含まれています。 これらの顆粒はエオシンで簡単に真っ赤に染色され、酸に溶けますが、アルカリには耐性があります。これらの細胞には、酵素(酸性ホスファターゼ、デヒドロゲナーゼ、ジペプチダーゼ)だけでなく、大量の亜鉛が含まれています。細胞パネス細胞は、これらの細胞によるリゾザイムの生成に関連する抗菌機能を実行し、細菌や原生動物の細胞壁を破壊します。これらの細胞は、微生物の活発な貪食が可能です。これらの特性により、パネス細胞は腸内細菌叢を調節します多くの病気でこれらの細胞の数は減少します近年これらの細胞でIgAとIgGが発見されましたさらにこれらの細胞はジペプチドをアミノ酸に分解するジペプチダーゼを生成しますそれらの分泌物が、カイムに含まれる塩酸を中和すること。
内分泌細胞拡散に属する 内分泌系。 すべての内分泌細胞が特徴づけられます
o分泌顆粒の核の下の基底部に存在するため、それらは基底顆粒と呼ばれます。 頂端表面には微絨毛があり、これには明らかに、pHの変化または胃の粥状液にアミノ酸がないことに反応する受容体が含まれています。 内分泌細胞は主にパラクリンです。 それらは、細胞の基底および基底外側表面を介して細胞間空間に分泌し、隣接する細胞に直接影響を及ぼします。 神経終末、平滑筋細胞、血管壁。 これらの細胞のホルモンの一部は血液中に分泌されます。
V 小腸最も一般的なのは次の内分泌細胞です:EC細胞(セロトニン、モチリン、物質Pを分泌)、A細胞(エンテログルカゴンを産生)、S細胞(セクレチンを産生)、I細胞(コレシストキニンを産生)、G細胞(ガストリンを産生)、D -細胞(ソマトスタチンを産生する)、D1-細胞(血管作用性腸ポリペプチドを分泌する)。 びまん性内分泌系の細胞は小腸に不均一に分布しています。それらの最大数は十二指腸の壁に見られます。 したがって、十二指腸では、100陰窩あたり150個の内分泌細胞があり、空腸と回腸には60個の細胞しかありません。
ボーダレスまたはボーダレスセル地下室の下部にあります。 彼らはしばしば有糸分裂を示します。 現代の概念によれば、フチなし細胞は低分化細胞であり、腸上皮の幹細胞として機能します。
自分の粘膜層ゆるい未成形から構築 結合組織。 この層は絨毛の大部分を構成します;陰窩の間に薄い層の形であります。 ここの結合組織は多くの細網線維と細網細胞を含み、非常に緩いです。 この層では、上皮の下の絨毛に血管叢があり、絨毛の中央にリンパ毛細血管があります。 物質はこれらの血管に入り、腸で吸収され、粘膜固有層の上皮と結合組織、および毛細血管壁を通って輸送されます。 タンパク質と炭水化物の加水分解生成物は毛細血管に吸収され、脂肪はリンパ毛細血管に吸収されます。
多数のリンパ球が粘膜の独自の層に位置しており、粘膜は単独で存在するか、単一の孤立したまたはグループ化されたリンパ濾胞の形でクラスターを形成します。 大きなリンパ球の蓄積はパイエル板と呼ばれます。 リンパ濾胞は粘膜下組織にも浸透する可能性があります。 ペイロフのプラークは主に回腸にあり、小腸の他の部分にはあまり見られません。 パイエル板の含有量が最も高いのは思春期(約250)で、成人ではその数が安定し、老年期(50-100)で急激に減少します。 t.propriaにあるすべてのリンパ球(単独およびグループ化)は、最大40%の免疫細胞(エフェクター)を含む腸関連リンパ系を形成します。 また、現在、小腸壁のリンパ組織はファブリキウスの袋に相当します。 好酸球、好中球、形質細胞、その他の細胞要素は、固有層に絶えず見られます。
粘膜の筋層(筋層)平滑筋細胞の2つの層で構成されています:内側の円形と外側の縦方向。 内層から、単一の筋細胞が絨毛の厚さに浸透し、絨毛の収縮と、腸から吸収された生成物が豊富な血液とリンパ液の押し出しに寄与します。 このような収縮は1分間に数回発生します。
粘膜下組織それは、多数の弾性繊維を含む緩い、形成されていない結合組織から作られています。 これが強力な血管(静脈)神経叢と神経叢(粘膜下組織またはマイスナー神経叢)です。 粘膜下組織の十二指腸には多数あります 十二指腸(ブルンナー腺)。 これらの腺は複雑で、分岐しており、肺胞-管状の構造をしています。 それらの末端部分は、平らな基底に横たわる核、発達した分泌装置、および頂端に分泌顆粒を有する立方体または円筒形の細胞で裏打ちされている。 それらの排泄管は陰窩に通じているか、または絨毛の基部で直接腸腔に通じています。 粘液細胞には、びまん性内分泌系に属する内分泌細胞が含まれています:Ec、G、D、S-細胞。 カンビア細胞は管の口にあるため、腺細胞の再生は管から末端部分に向かって起こります。 十二指腸腺の秘密には粘液が含まれています。粘液はアルカリ反応を起こし、それによって粘膜を機械的および化学的損傷から保護します。 これらの腺の秘密には、殺菌効果のあるリゾチーム、上皮細胞の増殖を刺激して胃での塩酸の分泌を阻害するウロガストロン、および酵素(ジペプチダーゼ、アミラーゼ、トリプシノーゲンをトリプシンに変換するエンテロキナーゼ)が含まれています。 一般的に、十二指腸腺の秘密は消化機能を果たし、加水分解と吸収のプロセスに関与します。
筋膜それは平滑筋組織で構成されており、内側の円形と外側の縦方向の2つの層を形成しています。 これらの層は、筋肉間(アウアーバッハ)神経叢が存在する、ゆるい、形成されていない結合組織の薄い層によって分離されています。 筋肉膜のために、長さに沿った小腸の壁の局所的および蠕動性収縮が実行されます。
漿膜は腹膜の内臓シートであり、上に中皮で覆われた、緩い、形成されていない結合組織の薄い層で構成されています。 漿膜には常に多数の弾性繊維があります。
小児期の小腸の構造組織の特徴。 新生児の粘膜が薄くなり、浮き彫りが滑らかになります(絨毛や陰窩の数が少ない)。 思春期までに、絨毛と襞の数が増え、最大値に達します。 陰窩は大人のものより深いです。 表面からの粘膜は上皮で覆われており、その特徴的な特徴は、陰窩の底だけでなく絨毛の表面にもある好酸性の顆粒を有する細胞の含有量が高いことです。 粘膜は、豊富な血管新生と高い透過性を特徴とし、毒素や微生物の血液への吸収と中毒の発症に適した条件を作り出します。 反応性中心を有するリンパ濾胞は、新生児期の終わりに向かってのみ形成されます。 粘膜下神経叢は未成熟であり、神経芽細胞を含んでいます。 十二指腸では、腺は少なく、小さく、枝分かれしていません。 新生児の筋層が薄くなります。 小腸の最終的な構造形成は、4〜5年でのみ発生します。
胃腺の上皮は、いくつかの細胞のディファレンからなる高度に特殊化された組織であり、形成層は、腺の首の領域の分化が不十分な上皮細胞です。 これらの細胞は、N-チミジンの導入によって集中的に標識され、しばしば有糸分裂によって分裂し、胃粘膜の表面上皮と胃腺の上皮の両方の形成層を構成します。 したがって、新たに出現する細胞の分化と移動は、表面上皮に向かって、そして腺の深さに向かって、2つの方向に進みます。 胃の上皮の細胞の更新は1〜3日で起こります。
高度に特殊化された細胞は、自分自身を更新するのがはるかに遅い 上皮胃腺。
主要な外分泌細胞プロ酵素ペプシノーゲンを生成します。これは、酸性環境では、胃液の主成分であるペプシンの活性型に変わります。 外分泌細胞は、角柱状の形状、よく発達した顆粒状小胞体、分泌性チモーゲン顆粒を伴う好塩基性細胞質を有する。
壁側外分泌細胞-主な外分泌細胞および粘液細胞から外側に向かって腺の壁に位置する、大きく、丸みを帯びた、または不規則な角度の細胞。 細胞の細胞質は急激に好酸性です。 それは多数のミトコンドリアを含んでいます。 核は細胞の中心部にあります。 細胞質には、細胞間尿細管に入る細胞内分泌尿細管のシステムがあります。 多数の微絨毛が細胞内細管の内腔に突き出ています。 塩酸を形成するHおよびClイオンは、分泌細管を介して細胞からその頂端面まで除去されます。
壁細胞それらはまた、小腸でのビタミンBi2の吸収に必要な城の内部因子を分泌します。
粘液細胞-基底部に移動した、軽い細胞質と圧縮された核を持つ角柱状の粘液細胞。 電子顕微鏡は、粘液細胞の頂端部に多数の分泌顆粒を明らかにします。 粘液細胞は腺の主要部分、主に自分の腺の体にあります。 細胞の機能は粘液の産生です。
胃の内分泌細胞いくつかのセルラーディファレンで表され、その名前には文字の略語が使用されます(EC、ECL、G、P、D、Aなど)。 これらの細胞はすべて、他の上皮細胞よりも軽い細胞質を特徴としています。 内分泌細胞の特徴は、細胞質に分泌顆粒が存在することです。 顆粒は硝酸銀を還元できるため、これらの細胞は好銀性と呼ばれます。 それらはまた、重クロム酸カリウムで強く染色されます。これが、内分泌細胞の別名である腸クロム親和性の理由です。
分泌顆粒の構造に基づいて、またそれらの生化学的および機能的特性を考慮に入れて、内分泌細胞はいくつかのタイプに分類されます。
EC細胞最も多くは、主な外分泌細胞の間の腺の体と底に位置し、セロトニンとメラトニンを分泌します。 セロトニンは、主要な外分泌細胞と粘液細胞の分泌活性を刺激します。 メラトニンは、光の周期に応じて分泌細胞の機能的活動の生体リズムの調節に関与しています。
ECL細胞壁側外分泌細胞に作用し、塩酸の生成を調節するヒスタミンを生成します。
G細胞ガストリン生成と呼ばれます。 V 大量にそれらは胃の幽門腺に見られます。 ガストリンは、ペプシノーゲンと塩酸の生成の増加を伴う、主および壁側外分泌細胞の活動を刺激します。 との人々で 過酸性胃液では、G細胞の数とその機能亢進が増加しています。 G細胞がエンケファリンを生成するという証拠があります。エンケファリンは、脳で最初に発見され、痛みの調節に関与するモルヒネのような物質です。
Pセル胆嚢の平滑筋組織の収縮を促進するボンベシンを分泌し、壁側外分泌細胞による塩酸の放出を刺激します。
Dセル成長ホルモン阻害剤であるソマトスタチンを産生します。 タンパク質合成を阻害します。
VIPセル拡張する血管腸ペプチドを生成します 血管と削減 動脈圧。 このペプチドはまた、膵島の細胞によるホルモンの分泌を刺激します。
Aセル膵島のグルカゴンA細胞と同様に、グリコーゲンをグルコースに分解するエンテログルカゴンを合成します。
多くの 内分泌細胞分泌顆粒は基底部にあります。 顆粒の内容物は粘膜の固有層に分泌され、次に毛細血管に入ります。
粘膜筋板滑らかな筋細胞の3つの層によって形成されます。
胃壁の粘膜下組織血管叢と神経叢を伴う緩い線維性結合組織によって表されます。
胃の筋層平滑筋組織の3つの層で構成されています:外側の縦方向、中央の円形、内側の筋肉束の斜め方向。 幽門領域の中間層は厚くなり、幽門括約筋を形成します。 胃の漿膜は、表面的に横たわっている中皮によって形成され、その基礎は緩い線維性結合組織です。
胃の壁に粘膜下神経叢、筋肉間神経叢、漿液性神経叢があります。 筋肉間神経叢の神経節では、第1型の栄養ニューロンが優勢であり、胃の幽門領域では、第2型のニューロンがより多く存在します。 迷走神経および境界交感神経幹からの導体は神経叢に行きます。 迷走神経の興奮は胃液の分泌を刺激し、交感神経の興奮は逆に胃液の分泌を抑制します。
腸
腸は、大腸と小腸の2つの部分で構成されています。 それは、消化管の上にある部分で始まった食物の消化のプロセスを続けます。
小腸の長さは4〜5 mに達し、十二指腸(30 cm)、空腸(2 m)、回腸(3 m)の3つのセクションで構成され、次の機能を実行します。
1.消化器系:それ自身の酵素と膵臓酵素の影響下で、タンパク質(エンテロキナーゼ、トリプシン、エリプシン)、脂質(リパーゼ)、炭水化物(アミラーゼ、マルターゼ、スクラーゼ、ラクターゼ)およびヌクレオタンパク質(ヌクレアーゼ)が切断されます。
2.分泌物:腸液の分泌:水、酵素(ジペプチダーゼ)、ミネラル、粘液で構成されています。
3.栄養素の崩壊生成物の吸収。 吸収面を大きくするために、腸粘膜には絨毛があり、上皮細胞には刷子縁があります。
4.粥状液を混合し、それを腸の下部に移動させることからなる運動排出機能。
5.バリア保護。 後者の基礎は、分泌された粘液、外皮上皮、リンパ濾胞および免疫担当細胞です。
6.内分泌:内分泌細胞によるAPUDシステムの生成 さまざまなホルモン、腸液の分泌、血液供給、運動性などの調節を提供します。
構造。 小腸の壁は、粘液、筋肉、漿膜の3つの膜で形成されています。 粘膜は、上皮、固有層、筋層、および粘膜下組織で構成されています。 小腸の粘膜のレリーフの特徴は、小腸の総表面を増加させる円形のひだ、絨毛および陰窩の存在であり、それはその主な機能の実行に貢献します。
ケルクリング皺は、粘膜が腸腔に突き出ている部分です。
腸絨毛は、上皮で覆われた固有層の腸管腔への指のような突起です。 上皮の基底膜の下にある緩い線維性結合組織には、毛細血管の密なネットワークがあり、絨毛の中心にはリンパ毛細血管があります。 絨毛の間質には、絨毛の動きを確実にし、血液やリンパ液に吸収された食物分解産物を動かすプロセスに寄与する単一の滑らかな筋細胞もあります。 絨毛の表面は、円筒形の境界上皮の単層で覆われています。 3種類のセルで構成されています。 境界上皮細胞、杯細胞および内分泌。
境界上皮細胞は最も多く、構造の顕著な極性によって区別されます。 頂端面には、高さ約1 µm、直径0.1 µmの微絨毛が含まれています。 細胞内のそれらの数は2〜3千に達し、一緒になって境界を形成し、粘膜の吸収面を30〜40倍に増やします。 微絨毛の表面には、リポタンパク質と糖タンパク質に代表される糖衣があります。 微絨毛の膜と糖衣には、頭頂部と膜の消化に関与するだけでなく、結果として生じるモノマー(グルコース、アミノ酸)の吸収機能に関与する多数の酵素が含まれています。脂肪は食作用によって、または次の形で吸収されます。それらの構成成分-グリセロールとアミノ酸。 細胞質には、発達した小胞体、ゴルジ複合体、ミトコンドリア、およびリソソーム装置があります。 隣接する上皮細胞の頂端部の膜は、腸腔からの物質の浸透を防ぐ閉鎖板を形成します。
絨毛の杯細胞は、辺縁系の上皮細胞の間に単独で位置し、粘液分泌物を生成します。 それらはゴブレットの形をしており、その拡張された頂端部には粘液を含む分泌顆粒があります。 後者は粘膜の表面に目立ち、それを保湿し、腸に沿った粥状液の動きに貢献します。
杯細胞のような内分泌細胞は、境界上皮細胞の間に単独で散在しています。
陰窩は、固有層への上皮の管状内殖です。 それらの内腔は、隣接する絨毛の基部の間で開きます。 小腸では約1億5000万個です。陰窩の上皮細胞の中には、上記に加えて、絨毛の上皮(境界、ゴブレット、内分泌)に境界のない細胞と好酸性の顆粒を持った細胞があります。 (パネート細胞)。 これらの細胞はすべて、絨毛の細胞とは対照的に、高さが低く、線条の境界が薄く、細胞質がより好塩基性です。 ボーダレス細胞は、陰窩および絨毛の上皮の生理学的再生の源である低分化細胞の集団です。 それらが増殖して分化するにつれて、これらの細胞は陰窩の基部から絨毛の上部に移動し、細胞を完全な細胞周期に置き換えます。 したがって、絨毛の上皮細胞の完全な交換は、3〜5日で行われます。
パネート細胞は、陰窩の基部にグループで配置されています。 これらは角柱状の細胞であり、その頂端部には大きな好酸性(ガマ腫。細胞核と細胞質網状組織が基底極に移動しています。これらの細胞の分泌産物はジペプチダーゼ(ジペプチドをアミノ酸に分解する酵素)です。これらの細胞はまた、破壊するリゾチームを生成すると考えられています 細胞壁バクテリア。
内分泌細胞。 EC細胞はEC1細胞とEC2細胞に分けられます。 前者はホルモンのメラトニンとセロトニンを生成し、後者はモチリンとサブスタンスPです。モチリンは絨毛の運動活動を刺激します。サブスタンスPは粘膜への血液供給を調節します。 セロトニンは粘液の形成を刺激し、血液循環と腸の運動性に影響を与えます。 メラトニンは、時間帯(照度)に応じて機能するバイオリズムに関与しています。
ECL細胞はヒスタミンを産生し、これが胃の主細胞と壁細胞を刺激します。
A細胞はエンテログルカゴンを産生します。エンテログルカゴンは血糖値を上昇させ、胃の外皮上皮による粘液産生を刺激します。
G細胞はガストリンを生成し、胃の主細胞と壁細胞を刺激します。
D細胞はソマトスタチンを形成し、D 1細胞は血管腸ポリペプチド(VIP)を形成します。 ソマトスタチンは腺の分泌と細胞分裂を阻害し、VIP-平滑筋を弛緩させ、血管を拡張し、血圧を下げます。
S細胞はセクレチンを産生し、膵臓による重炭酸塩の分泌を刺激します。
I細胞は膵臓を刺激するパンクレオチミンと胆嚢を収縮させるコレシストキニンを産生します。
小腸の粘膜の固有層は、多数の細網および弾性線維、血およびリンパ毛細血管の神経叢を伴う、緩い、形成されていない結合組織によって形成されます。 リンパ濾胞もあります。 回腸方向の後者の数が増加します。 彼らは大人よりも子供によく見られます。 リンパ球に加えて、形質細胞と好酸球が固有層に見られます。 粘膜の筋板は発達が不十分であり、内側の円形および外側の縦方向の筋層によって表されます。
粘膜下組織は、緩い、形成されていない結合組織によって形成され、血液の神経叢と リンパ管および神経叢(粘膜下組織)。 V 十二指腸十二指腸(ブルンナー)腺の末端部分があります。 構造上、これらは複雑な分岐管状腺です。 末端切片は、粘液細胞といくつかの頂端顆粒細胞(濃厚赤血球)によって形成されます。
筋肉のコートは、平滑筋組織の2つの層で構成されています。内側の円形と外側の縦方向です。 両方の層はらせん状です。 結合組織の層の層の間には、蠕動運動を提供する筋肉間血管叢と神経叢があります。
漿膜は、中皮で覆われた、下にある結合組織の基部によって形成されます。
小腸での消化と吸収のプロセスの組織生理学
空洞、頭頂葉、膜および細胞内消化を区別します。 消化は酵素の助けを借りて行われます 消化腺(唾液、胃、膵臓、腸)。 頭頂部の消化は、絨毛を覆う粘液層の食物を分解します。 膜消化は、絨毛腸細胞の刷子縁の糖衣で起こります。 細胞内-食作用および飲作用による腸細胞内。
小腸での消化と吸収の主な構造は絨毛です。 後者は、ストロマに滑らかな筋細胞が存在するため、収縮してポンプとして機能します。
炭水化物は単糖(グルコース)に分裂した後に吸収され、キャリアタンパク質の助けを借りて微絨毛膜を通して吸収されます。 タンパク質は、アミノ酸に分解した後、同様の方法で吸収されます。 胆汁酸の影響下にある腸腔内の脂肪は乳化されます(脂肪滴までは0.5ミクロン以下)。 小さな脂肪滴は腸細胞の糖衣に吸着され、リパーゼの影響下で脂肪はグリセロールと脂肪酸に分解されます。 脂肪酸はコレステロールと結合し(エステル化)、その後細胞に容易に浸透します。 腸細胞の無顆粒細胞質網状組織では、脂肪酸が吸収されたグリセロールと再結合して体特異的な脂肪を形成します。 タンパク質と炭水化物の成分は、顆粒状の細胞質網状組織とゴルジ複合体の後者に順次付着しています。 ゴルジ複合体で得られた複合化合物は、膜(カイロミクロン)に囲まれた顆粒に形成されます。 それらは80%以上のトリグリセリド、コレステロール(8%)、リン脂質(7%)、タンパク質(2%)などを含んでいます。それらは絨毛のリンパ毛細血管に入ります。
絨毛の尾側方向の小腸の構造的特徴は長くなり、薄くなり、上皮の杯細胞の数が増加します。 固有層では、リンパ濾胞の数が増加し、それらはクラスターを形成します-パイエル板。空腸と回腸には十二指腸腺はありません。
人間の胃が機能する原理を理解するには、細胞の構造と分類など、すべての詳細を分析する価値があります。 それらは胃液の重要な成分の1つである塩酸を生成します。
胃の形と大きさ
中空です 筋肉器官、いくつかの部分で構成され、消化機能を実行します。 違反した場合、 臨床症状。 胃は消化管の広い部分で、レトルトのような形をしており、十二指腸と食道の間にあります。
人体の位置、膨満感、機能状態、顔色によって変化が起こるため、恒久的な形ではありません。
たとえば、短気型の体型の人では、胃は角のように見え、ほぼ横方向に位置しています。 ドリコモルフィック型に属する人では、この器官は細長いストッキングのように見え、ほぼ垂直に配置されており、下部では右に鋭く曲がっています。 人が中型の体型をしている場合、胃はフックに似ています-その長い部分は上から下、そして左から右に向けられています。
空腹時の容量は約500mlです。 胃がいっぱいでない場合は、長さ14〜30cm、幅10〜16cmです。臓器の容量は1.5〜2.5リットルですが、4リットルになることもあります。
男性は女性よりもお腹が大きいことを覚えておいてください。 そして子供では、この器官は最も少ないです。 70キログラムの人では、胃の重さは平均150グラムです。
サイズの増加はストレスを引き起こす可能性があります、 慢性疲労, 炎症性疾患不規則な食生活。 胃がいっぱいになると食べ物の消化が遅くなるので、1つのモードで少しずつ食べる方が良いでしょう。 食べ過ぎは許されません。少し空腹感を残すことが望ましいです。
液体と一緒に消費される食物の量は、胃の2/3以下を占めるべきです。 この場合、伸びません。 ただし、食品の量に加えて、その組成を考慮する価値があります-有害で脂肪の多い食品、ガス形成食品が占める 大きなプロット食べ過ぎの感覚を引き起こします。
壁細胞
壁細胞はピラミッドや球のような形をしています。 彼らには国境を越えた理由があります 外面胃腺の体。 これらの細胞には、多数の楕円形ミトコンドリア、ゴルジ複合体、顆粒状ネットワークの短い槽、無顆粒状ネットワークの細管、遊離リボソームおよびリソソームが含まれていることが起こります。
腺細胞とも呼ばれる細胞の強い好酸性は、多くのミトコンドリアと滑らかな膜の蓄積の結果です。 それらは複合体とデスモソームによって近くの細胞に接続されています。
壁細胞は胃底腺の外側にあります。 男性の場合、その数は0.96から12.6億まで、女性の場合は0.69から0.91までさまざまです。 これらの細胞のうち10億個は、1時間以内に約23ミリモルの塩酸を分泌します。 男性の塩酸分泌の最大量は22〜29ミリモル、女性の塩酸分泌量は16〜21ミリモルです。
胃の壁細胞による塩酸の分泌は、水素イオンの膜貫通移動によって行われ、 プロトンポンプ。 このプロセスの最も重要な刺激物質は、ヒスタミン、アセチルコリン、ガストリンです。 それらは、胃の壁細胞の基底膜にある細胞受容体を介して作用します(これは壁細胞の別名です)。 受容体への曝露の結果として、アデノシン一リン酸とカルシウムの濃度が増加します。 そして塩酸の分泌の抑制剤はプロスタグランジンおよびソマトスタチンです。
壁細胞はまた、胃でのB12吸収と回腸でのその吸収に関与する糖タンパク質を分泌します。 赤芽球はこのビタミンなしでは成熟型に分化する能力がないため、これは非常に重要です。
悪意のある細胞
なぜ有益な細胞のいずれかが突然悪性になることができますか? 統計によると、それは最も一般的な腫瘍です。 がん患者の総数による死亡者数は38.48%です。
このような細胞は、以下の要因の影響の結果として形成されます。
- 揚げ物、脂肪、缶詰、辛い食べ物の乱用。
- 喫煙またはアルコール依存症。
- 、、などの慢性疾患。
- 遺伝的素因。
- 憲法の特徴。
- ホルモン活性。
- 長期投薬。
- 放射線の影響。
上級の専門家でさえ、胃がんの診断は簡単ではないと言うでしょう。 プロセスが非常に遅く、症状が他の病気と類似しているという事実のために、腫瘍を認識することは非常に困難です。
対症療法は特定することです 特徴的な症状胃または十二指腸の他の病理に存在します。 それらの範囲は広いので、腫瘍学についてすぐに話す価値はありません。それは患者を怖がらせるだけです。 次のような診断方法に頼る必要があります。 実験室研究、 CTスキャン。
このような有害な細胞の形成を防ぐために、観察する必要があります 健康的な生活様式人生、維持 適切な栄養。 胃を保護することができる食品はたくさんあります。 しかし、多くの場合、人々はそのような予防策について考えず、不適切に食べます-彼らは外出先で食べ、食べ過ぎ、脂肪分の多い食べ物を乱用します。
対照的に、抗ガン要素を含む野菜や果物があります-これらはブロッコリーです、 カリフラワー、大豆、玉ねぎ、にんにく、ナッツ、中国と日本のキノコ、魚、卵、トマト、柑橘系の果物。
胃はまた、角柱状、子宮頸部、粘液、主な内分泌細胞で構成されています。 それらのすべては体の正常な機能に責任があり、各タイプは特定の機能に責任があります。 頭頂骨は、腺の体の中で優勢であり、主要なものよりも大きいという理由で際立っています。
胃の主な機能は、 一次処理製品。 消化は、消化管の他の臓器との相互作用によって起こります。
胃の解剖学についての有用なビデオ
| 胃腺 | 分泌細胞 | 分泌物 |
| 基本的 | 主要 | ペプシノーゲン |
| オーバーレイ(または頭頂) | HC1 | |
| 追加 | 粘膜ムコ多糖類、城の内因子。 食物摂取とともに分泌が増加する | |
| 心臓 | 追加(主細胞と壁細胞はほとんどありません) | スライム |
| 幽門 | に似た主なもの | ペプシノーゲン |
| 眼底細胞 | 弱アルカリ性と | |
| 腺 | ねばねば、ぬるぬる。 | |
| 追加 | 分泌は食物摂取によって刺激されません | |
| 外皮-上皮- | セルは円筒形です | 粘液と液体が弱い |
| 細胞 | その上皮 | 局所反応 |
哺乳類の純粋な胃液は無色です 透明な液体酸反応(pH 0.8 ... 1.0); 塩酸(HCl)と無機イオン(カリウム、ナトリウム、アンモニウム、マグネシウム、カルシウムカチオン、塩化物アニオン、少量の硫酸塩、リン酸塩、重炭酸塩)が含まれています。 有機物質は、タンパク質化合物、乳酸、ブドウ糖、クレアチンリン酸、尿素、尿酸によって表されます。 タンパク質化合物は主にタンパク質分解酵素と脂肪分解酵素であり、そのうちペプシンが胃の消化に最も重要な役割を果たします。
ペプシンはタンパク質を高分子化合物(ポリペプチド(アルブモースとペプトン))に加水分解します。 ペプシンは、胃粘膜によって不活性なペプシノーゲンの形で生成され、酸性環境では、それらの活性な形であるペプシンに変わります。 既知の8 ... 11種類のペプシ-
新しい、機能的な機能に応じていくつかのグループに細分されます。
ペプシンA-酵素のグループ; オプチウムpH1.5 ... 2.0;
ペプシンC(ガストリキシン、胃カテプシン); 最適pH3.2 ... 3.5;
ペプシンB(パラペプシン、ゼラチナーゼ)-ゼラチンを液化し、結合組織タンパク質を分解します。 5.6までの最適pH;
ペプシンD(レンニン、キモシン)-乳タンパク質のカゼインをカゼインに変換します。カゼインはカルシウム塩として沈殿し、緩い血餅を形成します。 キモシンはカルシウムイオンによって活性化されます。 ミルク期間中に動物の胃の中で大量に形成されます。 カゼインとそれに吸着した乳化乳脂肪は胃に保持され、消化しやすいアルブミン、グロブリン、乳糖を含む乳清が腸に排出されます。
胃液リパーゼは脂肪に対する加水分解効果が弱く、乳脂肪などの乳化脂肪を最大限に分解します。
塩酸は胃液の重要な成分です。 峡部と胃の上半身にある壁細胞によって生成されます。 塩酸は、胃腺と膵臓腺の分泌の調節に関与し、ガストリンとセクレチンの形成を刺激し、ペプシノーゲンのペプシンへの変換を促進し、ペプシンの作用に最適なpHを作り出し、タンパク質の変性と腫れを引き起こします。胃から十二指腸への食物の通過を促進し、十二指腸粘膜のエンテロキナーゼ酵素腸細胞の分泌を刺激し、胃の運動活動を刺激し、幽門反射の実施に参加し、殺菌効果を有する。
塩酸の分泌はcAMPに依存するプロセスです。 カルシウムイオンは、塩酸分泌システムの機能に必要です。 酸生成細胞の働きは、H +イオンの喪失と細胞内のOH-イオンの蓄積を伴い、細胞構造に損傷を与える可能性があります。 それらの中和反応は、胃の炭酸脱水酵素によって活性化されます。 この場合に形成された重炭酸イオンは血液中に排泄され、C1〜イオンが代わりに細胞に入ります。 塩酸の分泌過程における主要な役割は、細胞のATPアーゼのシステムによって果たされます。 NA + / K + --ATPzaは、血液からのNa +と引き換えにK +を輸送し、H + / K + --ATPzaは、胃液に排出されるH +イオンと引き換えに一次秘密からK +を輸送します。
胃液には少量の粘液が含まれています。 粘液(ムチン)は、追加の細胞(粘液細胞)と胃腺の表面上皮の細胞の分泌産物です。 それは中性ムコ多糖類、シアロムシン、グリコサミノグリカンで構成されています-
コプロテインとグリカン。 ムチンは胃粘膜を包み込み、外因性因子の損傷作用を防ぎます。 粘膜細胞は重炭酸塩も生成し、ムチンと一緒になって、塩酸とペプシンの影響下で粘膜を自己消化(自己消化)から保護する粘膜-重炭酸塩バリアを形成します。 胃壁へのペプシンの作用は、循環血液のアルカリ反応によっても防止されます。
胃液の分泌の調節。 V胃液分泌には、刺激性因子の影響の特徴に関連する3つの主要な段階があります。 胃の神経液性; 腸の体液性。
分泌の最初の段階である複雑な反射は、無条件および条件付きの反射メカニズムの複雑な複合体の結果です。 その始まりは、対応する分析装置の受容体に対する食物の種類と匂いの影響(条件付けされた刺激)、または食物による口腔の受容体の直接的な刺激(条件付けされていない刺激)に関連しています。 胃液の分泌は、食べてから1〜2分後に起こります。 I.P.パブロフは、この期間を「発火」と呼びました。これは、その後の胃と腸の消化プロセスがそれに依存しているためです。 塩酸と酵素が高濃度に含まれています。
複雑な反射期の存在は、食道切開(食道の切除)後に犬を使用する、いわゆる「想像上の摂食」を用いた実験で、IPパブロフによって説得力のある形で証明されました。 この場合、食道の端が引き出され、首の皮膚に縫い付けられました。 このように、犬に吸収された食物は、胃に入ることなく食道の上端から落ちました。 「架空の摂食」が始まってからしばらくすると、酸性度の高い胃液が大量に放出されました。
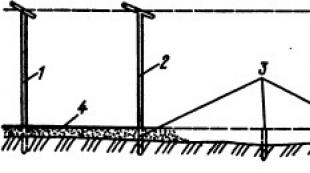
胃液分泌を研究するために、ハイデンハインは使用しました 外科的方法主胃の空洞からの小心室の隔離(図5.4)。 したがって、小心室から分泌されるジュースには食品の不純物はありませんでした。 しかし、この方法の主な欠点は、手術中の神経幹の切断による小心室の除神経です。 このような心室での胃液の分泌は、犬に餌を与えてから30〜40分後に始まりました。
IPパブロフは、小さな心室を切除するためのまったく新しい方法を提案しました。この方法では、その神経支配が妨げられることはありませんでした。 小心室の空洞は、迷走神経の枝の完全性を維持しながら、粘膜を犠牲にしてのみ大心室から隔離されました(図5.4を参照)。 パブロフ法に従って分離された小心室での胃液の分泌は、食事の1〜2分後に始まりました。
米。 5.4。 小さな分離スキーム
ハイデンハインによる心室 (A)と
I.P.パブロフ (B):
1 -孤立した心室; 2行のカット; 3 -迷走神経の枝; 4- I. P. Pavlovによると、大きな胃と孤立した心室の間の神経筋接続。 孤立した心室に供給する血管を伴う5-腸間膜
したがって、胃液分泌の第一段階の実施のための中枢神経系および胃の神経支配の役割が証明された。
口腔受容体からの求心性経路は唾液反射と同じです。 胃液分泌の神経中心は迷走神経の核にあります。 から 神経中枢延髄の胃腺への興奮は、迷走神経の分泌神経線維を介して伝達されます。 犬の両方の迷走神経が切断された場合、「想像上の摂食」は胃液の放出を引き起こしません。 胃腺、主に粘液細胞の分泌の調節における交感神経の関与は、実験的に証明されています。 交感神経が通過する太陽神経叢の除去 神経線維胃は、胃腺の分泌の急激な増加につながります。
胃液分泌の反射期は、第2期である神経液性と重なっています。 それは、餌の開始から30〜40分後に始まり、食塊による胃壁の機械的および化学的刺激を伴います。 胃液分泌の神経液性調節は、ホルモン、飼料の抽出物、栄養素の加水分解生成物などの生物学的に活性な物質の作用により行われます。 消化産物と食物の抽出物質は、胃の幽門部の血液に吸収され、血流とともに胃底腺に運ばれます。
刺激 食べ物の塊胃の壁は、胃腸管のホルモンの1つである粘膜の特殊な細胞による産生につながります- ガストリナ。ガストリンは胃の幽門部で不活性状態(プロガストリン)で生成され、塩酸の作用で活性物質に変換されます。 ガストリンは、次のような生物学的に活性な物質の放出を刺激します ヒスタミン。ガストリンとヒスタミンは、主に塩酸である胃液分泌に刺激効果があります。
消化管で合成された生物活性物質は、その粘膜の細胞の頂端膜の側面から直接作用する可能性があることに注意する必要があります。 同時に、それらは血液に吸収され、粘膜下膜および基底膜から壁内神経系を介して上皮細胞に作用することができます。
胃液分泌の第3段階- 腸の体液性-部分的に消化された食物昏睡が十二指腸に入ったときに始まります。 タンパク質加水分解の中間生成物がその粘膜に作用すると、ホルモンが放出されます モチリン、胃液分泌を刺激します。 十二指腸の粘膜と空腸の最初の部分で、ポリペプチドが形成されます- エンテロガストロン、その作用はガストリンに似ています。 消化産物(特にタンパク質)は、腸の血液に吸収され、胃腺を刺激し、ヒスタミンとガストリンの形成を増加させる可能性があります。
胃腺の分泌活動を刺激する物質に加えて、胃と腸で形成され、胃液分泌の阻害を引き起こします。 デリと エンテロロガストロン。これらの物質は両方ともポリペプチドです。 胃の幽門部にガストロンが形成され、胃底腺の分泌を抑制します。 エンテロガストロンは粘膜で合成されます 薄い部門脂肪、脂肪酸、塩酸、単糖にさらされたときの腸。 十二指腸の内容物のpHが4.0を下回った後、酸性粥状液はホルモンを生成し始めます セクレチン、胃液分泌を抑制します。
ホルモンはまた、胃液分泌を阻害する体液性因子に属します。 ブルボガストロン、胃抑制ポリペプチド(gip)、 コレシストキニン、血管作動性腸管ペプチド(VIP)。 さらに、脂肪のごく一部でさえ、胃底の細胞の分泌活性を急激に阻害します。
食物を構成する物質は、胃液分泌の適切な調節因子です。 同時に、胃の分泌装置はその質、量、食事に適応します。 肉食(犬)は胃液の酸性度と消化力を高めます。 タンパク質とその消化産物は顕著な樹液作用を示し、胃液の最大分泌は食事後2時間で起こります。 炭水化物食品は分泌をわずかに刺激します:食事後の最初の1時間で最大になります。 その後、分泌物が急激に低下し、長期間低レベルに保たれます。 炭水化物ダイエットは、ジュースの酸性度と消化力を低下させます。 脂肪は胃液分泌を阻害しますが、食後3時間の終わりまでに、分泌反応は最大に達します。
胃の運動活動。非活動状態(食物摂取不足)では、胃の筋肉は収縮した状態にあります。 食べることは胃の壁の反射弛緩につながり、それは胃の空洞への食物昏睡の沈着と胃液の輸送に貢献します。
胃壁の平滑筋は自発的な活動(自動化)が可能です。 彼らにとって適切な刺激物は、食物で胃の壁を伸ばすことです。 満腹の場合、収縮には主に2つのタイプがあります。強直性と蠕動性です。 強直性収縮は、縦筋層と斜筋層の波状の圧迫の形で現れます。 蠕動収縮は、収縮リングの波のような動きの形で強壮剤の背景に対して行われます。 それらは、不完全な輪状狭窄の形で胃の心臓部分から始まり、徐々に増加し、幽門括約筋に移動します。 収縮の輪の下で、筋肉部分の弛緩が起こります。
十二指腸の空洞への食物昏睡の動きは断続的であり、胃と十二指腸の機械受容器と化学受容器の刺激によって調節されます。 胃の機械受容器の刺激は避難を加速し、腸の刺激はそれを遅くします。
幽門反射は、胃(酸)と十二指腸(アルカリ性)の空洞における環境の異なる反応によるものです。 十二指腸に入ると酸性反応を示す粥状液の一部は、その化学受容器に非常に強い刺激作用を及ぼします。 その結果、幽門括約筋(閉塞具幽門反射)の円形筋が反射的に収縮し、内容物が完全に中和されるまで、粥状液の次の部分が十二指腸腔に入るのを防ぎます。 括約筋が閉じると、残りの胃内容物は胃の幽門部に戻されます。 そのようなダイナミクスは、胃の中で食物内容物と胃液の混合を確実にします。 胃の体内では、このような混合は起こりません。 十二指腸の内容物が中和された後、幽門括約筋が弛緩し、食物の次の部分が胃から腸に流れ込みます。
胃からの食物塊の排出速度は、多くの要因、主に食物含有量の量、組成、温度および反応、幽門括約筋の状態などに依存します。したがって、炭水化物が豊富な食物は、タンパク質が豊富なよりも胃から排出されます。 脂肪分の多い食品は最も遅い速度で避難します。 液体は胃に入った直後に腸に入り始めます。
胃の運動活動は、副交感神経(迷走神経)と交感神経(セリアック病)によって調節されています。 迷走神経、原則として、それを活性化し、セリアック病はそれを抑制します。 胃(および胃腸管全体)の神経支配の特徴は、その壁に大きな、いわゆる 壁内神経叢:筋肉の環状層と縦方向の層の間に局在する筋肉間(またはアウアーバッハ)神経叢、および粘膜と漿膜の間に位置する粘膜下(またはマイスナー)神経叢。 形態学的特徴、メディエーター組成、および同様の構造の生体電位の特徴。これらは子宮壁にも存在します。 膀胱平滑筋壁を備えた他の器官は、それらを特別な種類の植物として区別することを可能にします 神経系-交感神経系(交感神経および副交感神経とともに)。 このような壁内神経叢の神経節は完全に自律的な形成であり、独自の反射弧を持ち、完全に分散化されていても機能することができます。 無傷の生物では、中交感神経系の構造は、胃腸管のすべての機能の局所的(局所的)調節において重要です。
胃の筋肉を刺激する体液性因子は、ガストリン、ヒスタミン、モチリン、コレシストキニン、プロスタグランジンです。 抑制効果は、アドレナリン、ブルボガストロン、セクレチン、血管作動性腸管ペプチドおよび胃抑制ポリペプチドによって発揮されます。
空腹定期刊行物。 19世紀の終わりまで、食事の外であると信じられていました 消化管は「休息」の状態にあります。つまり、その腺は分泌されず、胃腸管は収縮しません。 しかし、すでにその時、人間と動物の空腹時に胃と腸の収縮の出現の証拠がありました。 IP Pavlovは、犬の長期実験で、胃の運動活動の期間を確立し、膵臓、腸の分泌、腸の運動性を同期的に増加させました。 彼はそのような胃の活動の中で、定期的に「仕事」と「休息」の期間を交互に繰り返し、それぞれ平均20分と80分を選びました。 周期的な活動の根本的な原因は生理的空腹の状態であるため、そのような収縮はと呼ばれます 空腹の定期刊行物。
胃の空腹活動のメカニズムは、視床下部の活性化、欠乏症に関連しています 栄養素血液、細胞内および細胞外液中。 視床下部は、脳の関与により、摂食行動を活性化します。 空腹と小腸の近位部分の空腹活動は、空腹感を悪化させ、動物の無意識の運動不安と人間の意識的な空腹感を引き起こします。
消化器の周期的な活動は、体に不要な物質の除去に貢献し、分泌はサポートします 通常のミクロフローラ腸、小腸へのミクロフローラの拡散を防ぎます。 消化液が定期的に放出されるため、粘膜、絨毛装置、腸細胞の刷子縁の正常な状態が維持されます。