女性の生殖器系の生理学。 月経周期。 健康な家族の形成と性感染症の予防の基礎としての性教育
子宮の形は洋ナシの形をしており、前後方向に平らになっています。 子宮の上部外側縁から、子宮(卵管)と卵巣が位置する広い子宮靭帯があります(図1)。 解剖学的に、子宮の底、体、子宮頸部は区別されます。
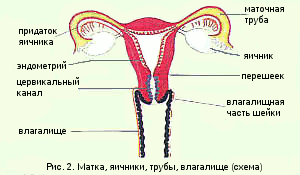
底は卵管の排出物の上にある子宮の部分です。 体は三角形の形をしており、峡部に向かって先細になっています。 子宮腔も三角形で、上部の角には卵管に通じる2つの穴があり、下部の角には峡部があります。これは子宮頸管の腔につながる狭窄です(図2)。
子宮頸部は、子宮の比較的狭い下部セグメントです。 女の子と女の子の場合は円錐形で、大人の女性の場合は円筒形です。 膣部分(portio vaginalis cervicis-ectocervix)、子宮頸管(canalis cervicalis uteri-endocervix)および峡部を割り当てます。 子宮頸部には2つの開口部があります。内部咽頭は体と子宮頸部の境界にある上部の開口部であり、外部咽頭は膣に通じる下部の開口部です。
子宮頸部の膣部分は丸い形をしており、その表面は滑らかで、中央には外咽頭があります。 もつ 未経産の女性形は小さく、丸い、または交差楕円形です(小魚の口)。 出産後、外咽頭は横スリットの形をとります。 子宮頸管は狭く、中央部が広くなっています。 前面と背面には2つの縦方向の隆起があり、そこから粘膜のひだ、手のひらのようなひだが斜めに分岐しています。 これらの地層は運河に奇妙な外観を与え、生命の木と呼ばれています。
膣は小さな骨盤にある筋肉弾性管であり、 上子宮頸部を覆い、下の方が生殖器の隙間に通じています。
子宮体の粘膜は、間質と、間質に成長して単純な管状腺を形成する単層円柱上皮で構成されています。 イスムスの粘膜は子宮体の粘膜に似ており、多数の結合組織細胞と単一の単純な分枝していない腺によって表されます。 体の粘膜と峡部は、月経周期の間に周期的な変化を受けます。
子宮頸部の壁は主にコラーゲン組織で構成されており、粘膜の間質には多くの弾性繊維が含まれています。 子宮頸管内腺は管状で分岐しており、構造が全体を通して同じであるため、真の腺とは見なされません。 腺には、アルカリ性反応を示す厚いガラス状の粘液の形で秘密が含まれています。 アルカリ反応は、精子の生存能力、子宮腔への移動の維持に貢献します。 排卵中の粘液の分泌が増加し、その秘密が子宮頸管を満たし、いわゆるクリステラープラグを形成します。これは、その殺菌特性により、微生物が子宮頸管と子宮腔に入るのを機械的に防ぎます。 腺が詰まり、粘液が蓄積し続けると、ナボット嚢胞が形成され、子宮頸部の表面に突き出る可能性があります。
運河の粘膜は、円柱状の粘液産生上皮によって表され、単一の繊毛細胞があり、それらの数は年齢とともに著しく減少します。
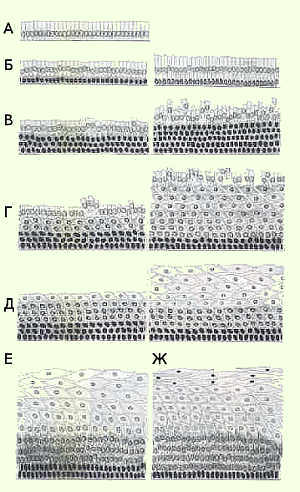
外頸部および膣の粘膜は、重層扁平上皮の角質化していない上皮で裏打ちされています。 通常、生殖年齢では、上皮は多くの列で構成され、通常、基底、中間、および表面の3つの層に分割されます。 細胞の下(基底)層だけが基底膜に接続されており、その中の細胞は一列に並んでいます。 基底細胞の上に位置する若い細胞の層で、いくつかの列(中間層の下部)で構成されています。 それらはパラベーサルと呼ばれます。 細胞サイズは成熟とともに増加します。 核のサイズが小さくなります(図3)
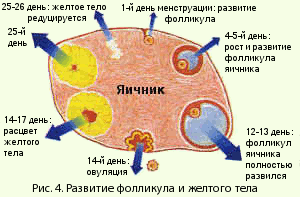
膣の重層扁平上皮と子宮頸部の膣部分の構造は、女性のホルモン状態、月経周期の段階に依存します。 月経周期には、月経、フォリキュリン(エストロゲン、増殖)、排卵、黄体(プロゲスチン、分泌)の4つの段階があります。 これらの段階は、視床下部-下垂体系(下垂体前葉)の性腺刺激ホルモンによって調節される卵子の成熟に関連しています。 FSHの作用下で、卵胞は卵巣で成長し成熟します(図4)。
成長中の卵胞はエストロゲンホルモンを分泌し、その一定量がFSHの産生を阻害し、LHの分泌を刺激します。 LHはFSHと一緒に卵胞を排卵の準備をし、卵胞の破裂(排卵)後、妊娠が起こらなければ、黄体への変換に寄与します。
膣上皮はホルモンの影響を最も受けやすいため、ホルモンの細胞学的診断はこの上皮の組成の研究に基づいています(ホルモンの細胞学的診断を参照)。
運河の円筒形の上皮は、子宮頸部の膣部分の女の子、生殖年齢の女性、外咽頭のレベルにある、いわゆる接合部で平らになります。 ホルモンや他の影響を受けている関節ゾーンは、子宮頸部の膣部分に移動する可能性があります。 子宮頸部の膣部分の円柱上皮の出現は、異所性と呼ばれます。 膣の内容物の影響下で、異所性部位は生理学的変化、扁平上皮への化生を起こします(図6)。
 |
1-成熟した上皮。 表面には、ピクノティック核を持つ表層の成熟細胞 2-上皮は中間層に成熟し、表面には成熟した中間細胞があります 3-上皮は中間層に成熟し、表面には未成熟な中間細胞があります 4-5-上皮は、傍基底細胞の表面の傍基底層までのみ成熟します |
図5。 重層扁平上皮の成熟のさまざまな段階 |
|
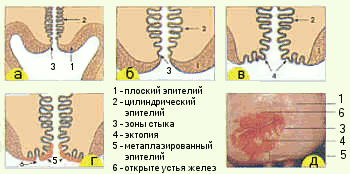 |
A-思春期前(関節ゾーンは子宮頸管にあります) B-C-思春期中(関節ゾーンは子宮頸部の膣部分に移動します) D-膣部分の円柱上皮が化生癌に置き換わっている D-経産婦の子宮頸部のコルポスコピー画像 |
図6。 円柱上皮の位置と子宮頸部の接合部領域 |
|
下垂体からのLHの作用下で形成された黄体はプロゲステロンを分泌します。 卵巣と副腎、卵巣とのホルモンの間には密接な関係があります 甲状腺.
エストロゲンは、重層扁平上皮の角質化していない上皮の表面細胞への完全な成熟を刺激します。 プロゲステロンは成熟を阻害し、それが大量に生成されると、細胞は中間層までしか成熟しません。 閉経後は、性ホルモンの産生が減少するため、上皮が萎縮します(図5)。
膣部分の円筒形の上皮は化生上皮に置き換えられます。 化生上皮の領域は、形質転換ゾーン、または形質転換ゾーンと呼ばれます。 生殖年齢の女性では、関節領域は通常、自然の関節領域の領域、変形ゾーンの領域、化生上皮によって表されます。 変換ゾーンは、子宮頸部の膣部分に配置することも、(全体的または部分的に)子宮頸管に挿入することもできます。 閉経後の女性では、関節ゾーンと変形ゾーンはほとんどの場合子宮頸管にあります。 形質転換帯は、腫瘍性変化を含む病理学的可能性の観点から最も危険です。
化生扁平上皮(図7)は、成熟した円柱状ではなく、円柱状のいわゆる予備細胞から発生します。 通常、予備細胞は通常、組織学的および細胞学的調製物には見られません。 予備細胞過形成は扁平上皮化生の最初の段階です。 生殖細胞の1つ、2つ、またはそれ以上の層が円柱状細胞の層の下に現れます。これは、明確な細胞境界のない扁平上皮の基底層の細胞に似ています。
成熟した化生上皮は、重層扁平非角化上皮に典型的なすべての層によって表される「天然の」扁平上皮と形態学的に実質的に区別がつかない。
 |
A-円柱上皮 B-円柱上皮の層の下に円柱下(予備)細胞の層が現れる B-予備細胞が増殖し、円筒形細胞が表面から剥離します D-未成熟扁平上皮化生の段階:予備細胞の明確な境界が決定され、多層扁平上皮非角質化上皮と同様に、細胞の3〜4層が徐々に形成されます D-成熟する扁平上皮化生の段階。 上皮層の表面に、小さな核を持つ中型の細胞 E-成熟扁平上皮化生の段階。 上皮層の表面の細胞は、扁平上皮の中間細胞に似ています G-成熟扁平上皮化生の段階。 上皮層の表面の細胞は、扁平上皮の表層の「天然の」細胞と実質的に区別がつかない。 |
図7。 扁平上皮化生の段階 |
|
(書類)
n1.doc
第2章女性生殖器系の解剖学と生理学
2.1。 女性の生殖器の解剖学
婦人向け 生殖システム骨盤腔内にある内性器(内性器)と骨盤外にある外性器(外性器)で構成されています。 内性器には、子宮、卵巣、卵管、および膣が含まれます。 外性器には、恥骨、大陰唇、小陰唇、膣の前庭、陰核などがあります。 骨盤腔は骨の構造に限定されています。 内性器と隣接する器官の底は、筋肉と筋膜からなる骨盤横隔膜です。
2.1.1. 女性の骨盤
骨盤(セクション2.1.3を参照)は、4つの骨で構成されています。2つの骨盤(または名前のない)骨、仙骨、および尾骨です。
16〜18歳までの骨盤骨(寛骨)は、腸骨、恥骨、坐骨の3つの骨で構成されています。 軟骨の骨化後、これらの骨は一緒に成長して骨盤(無名)の骨を形成します。
腸骨(os ilium)は、体と翼の2つの部分で構成されています。 体は、寛骨臼の形成に関与する骨の短くて厚い部分です。 翼は、内側が凹状で凸状の幅広のプレートです。 外面..。 翼の最も厚い自由端は腸骨稜(cristailiaca)を形成します。 前部では、尾根は上前腸骨棘、または突起(上前腸骨棘)から始まり、下は下前腸骨棘(下前腸骨棘)です。
後部の腸骨稜は、上後腸骨棘(上後腸骨棘)で終わり、その下に2番目の突起(下後腸骨棘(上後腸骨棘))があります。 腸骨の内面、翼から体への移行の領域には、弓状、境界線、または名前のない線を形成する尾根の突起があります(linea arcuata、s.terminalis、s.innominata )。 この線は仙骨から腸骨全体に渡り、正面では恥骨の上端に達します。
坐骨(os ischii)は、寛骨臼の形成に関与する体と、上部と下部の2つの枝で構成されています。 上部の枝は体から下に向かって坐骨神経痛(坐骨結節)で終わります。 下の枝の背面には突起があります-坐骨棘(spinaischiadica)。 下の枝は前方と上向きに向けられ、恥骨の下の枝に接続します。
恥骨(恥骨)は骨盤の前壁を形成します。 それは体と2つの枝で構成されています:上部(水平)と下部(下降)。 恥骨の短い体は寛骨臼の一部を形成し、下の枝は坐骨の対応する枝と結合します。
前の両方の恥骨の上部と下部の枝は、座りがちな恥骨の関節-シンフィシス(シンフィシス)を介して互いに接続されています。 両方の恥骨は、中間軟骨によって交感神経で接続されています。中間軟骨には、液体で満たされた小さなスリット空洞が含まれていることがよくあります。 恥骨の下部の枝は、恥骨弓と呼ばれる恥骨の下で角度を形成します。 恥骨と坐骨の接続枝は、かなり広い閉鎖孔開口部(閉鎖孔孔)を制限します。
仙骨(os sacrum)は、5つの椎骨で構成されています。 仙椎のサイズは下向きに減少するため、仙骨は円錐台の形をしています。 その広い部分(仙骨の基部)は上向きで、狭い部分(仙骨の頂点)は下向きです。 仙骨の後面は凸面であり、前面は凹面であり、仙骨腔を形成します。 仙骨の基部(I仙椎の表面)はV腰椎と関節でつながっています。 仙骨の基部の前面の中央に、突起が形成されます-仙骨岬(岬)。
尾骨(os coccygis)は、4〜5個の初歩的な尾骨の尾骨で構成され、下向きに先細になる小さな骨です。
骨盤の骨は、交感神経、仙腸関節、仙腸関節を介して接続されています。 軟骨層は骨盤の関節にあります。 骨盤の関節は強力な靭帯で補強されています。
2.1.2。 外性器と内性器
外性器(genitalia externa、s。vulva)は、総称「vulva」または「pudendum」を持ち、恥骨結合の下にあります(図2.1)。 これらには、恥骨、大陰唇および小陰唇、陰核、および前庭が含まれます。 膣の前夜に、尿道(尿道)の外部開口部と前庭の大きな腺(バルトリン腺)の管が開きます。
腹壁の境界領域である恥骨(mons pubis)は、恥骨結合と恥骨の前にある丸みを帯びた正中隆起です。 思春期後、それは髪で覆われるようになり、その皮下基部は、集中的な発達の結果として、脂肪パッドの形をとります。
大陰唇(labia pudendi majora)は、大量の脂肪組織と丸い子宮靭帯の線維性末端を含む皮膚の広い縦方向のひだです。 前部では、大陰唇の皮下脂肪組織が恥骨の脂肪パッドに入り、その後ろで坐骨直腸脂肪組織に接続されています。 思春期に達した後、大陰唇の外面の皮膚は色素沈着し、髪で覆われます。 大陰唇の皮膚には、汗と皮脂腺があります。 それらの内面は滑らかで、髪の毛で覆われておらず、皮脂腺で飽和しています。 前の大陰唇の関節は前交連と呼ばれ、後ろの陰唇交連、または 後部はんだ付け..。 陰唇の後交連の前の狭い空間は蝶形骨の舟と呼ばれます。
米。 2.1。 外性器。
1-恥骨; 2-前交連; 3-大 陰唇; 4-小陰唇; 5-膣の後壁; 6-膣の前庭の窩; 7-後交連(陰唇の交連); 8-肛門; 9-股間; 10-膣への入り口; 11-処女膜の自由端; 12-尿道の外部開口部; 13-陰核小帯; 14-クリトリス。
小さな陰唇(小陰唇)。 小陰唇と呼ばれるより小さな厚い皮膚のひだは、大陰唇の内側にあります。 大陰唇とは異なり、それらは毛で覆われておらず、皮下脂肪組織を含んでいません。 それらの間に膣の前庭があり、小陰唇が希釈されたときにのみ見えるようになります。 小陰唇がクリトリスと出会う前で、それらはクリトリスの周りで合流する2つの小さなひだに分かれます。 上部のひだは陰核の上で結合して陰核包皮を形成します。 下のひだは陰核の下側で結合して陰核小帯を形成します。
クリトリス(クリトリス)は、包皮の下の小陰唇の前端の間にあります。 それは男性の陰茎の海綿体の同族体であり、勃起することができます。 クリトリスの体は、繊維状の膜で囲まれた2つの海綿体で構成されています。 各海綿体は、対応する坐骨神経痛の内側縁に付着した椎弓根から始まります。 クリトリスは、靭帯サポートを使用して恥骨結合に取り付けられています。 クリトリスの体の自由端には、頭と呼ばれる小さな勃起組織の隆起があります。
前庭球(前庭球)は、小陰唇の深部に位置し、馬蹄形で前庭を覆う静脈叢です。 前庭の近く、各小陰唇の深部に沿って、前庭球と呼ばれる楕円形の勃起組織の塊があります。 それは静脈の密な神経叢によって表され、男性の陰茎の海綿状の体に対応します。 各球根は、泌尿生殖器横隔膜の下筋膜に付着し、球根スポンジ(球海綿体)の筋肉で覆われています。
膣の前庭(前庭膣)は小陰唇の間にあり、そこで膣は垂直のスリットの形で開きます。 開いた膣(いわゆる開口部)は、さまざまなサイズの線維組織(膜結節)の節に囲まれています。 小さな垂直スリットの形をした尿道の外部開口部は、正中線のクリトリスの頭から約2cm下の膣開口部の前にあります。 尿道の外部開口部の端は通常隆起しており、ひだを形成します。 尿道の外部開口部の両側には、尿道腺(尿道傍尿道)の管のミニチュア開口部があります。 膣口の後ろの小さなスペースは前庭窩と呼ばれます。 ここでは、両側で、大きな前庭腺、またはバルトリン腺(glandulae vestibulares majorus)の管が開いています。 腺はエンドウ豆の大きさの小さな小葉体であり、前庭球の後端に位置しています。 これらの腺は、多数の小さな前庭腺とともに、膣の前夜にも開いています。
内性器(性器内科)。 内性器には、膣、子宮、およびその付属肢である卵管と卵巣が含まれます(図2.2)。
膣(vaginas。Colpos)は、生殖器のスリットから子宮まで伸び、泌尿生殖器と骨盤の横隔膜を後方に傾斜して上向きに通過します(図2.3)。 膣の長さは約10cmです。これは主に骨盤腔にあり、そこで終わり、子宮頸部と合流します。 膣の前壁と後壁は通常、H字型の断面で下部で互いに接続されています。 子宮頸部の膣部分の周りに内腔がポケットまたは丸天井を形成するため、上部セクションは膣円蓋と呼ばれます。 膣は子宮に対して90°の角度にあるため、後壁は前部よりもかなり長く、後部円蓋は前部および外側の円蓋よりも深くなっています。 外側の膣壁は、子宮の心臓靭帯と骨盤横隔膜に取り付けられています。 壁は主に平滑筋と多くの弾性繊維を含む密な結合組織で構成されています。 外層には、動脈、神経、神経叢を伴う結合組織が含まれています。 粘膜には横方向と縦方向のひだがあります。 前部と後部の縦方向のひだは、ひだの柱と呼ばれます。 表面の重層扁平上皮は、月経周期に対応する周期的な変化を受けます。 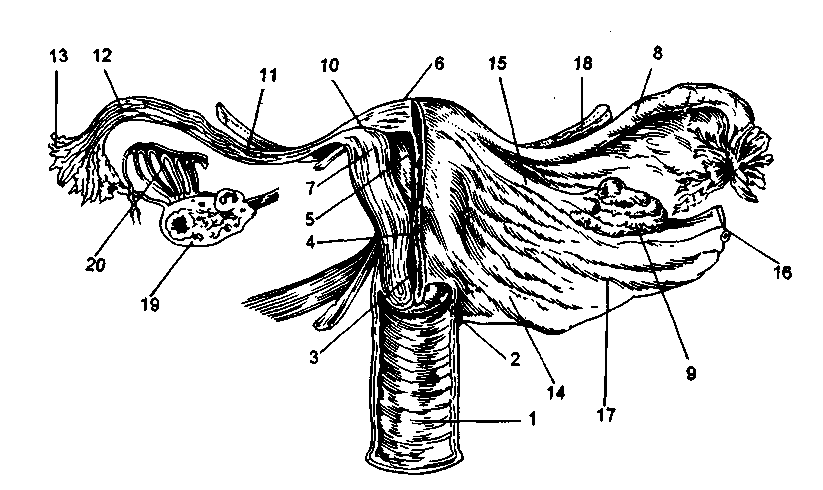
米。 2.2。 内臓。
1-膣; 2-子宮頸部の膣部分; 3-子宮頸管; 4-イスムス; 5-子宮腔; 6-子宮の底; 7-子宮の壁; 8-卵管; 9-卵巣; 10-チューブの間質部分。 11-パイプのisthmic部分。 12-パイプのアンプラ部分。 13-パイプのフィンブリア; 14-仙子宮靭帯; 15-卵巣の自身の靭帯; 16-漏斗靭帯; 17-広間膜; 18-丸い靭帯; 19-卵胞と黄体を伴う卵巣の切片。 20-蒸気タンク。
膣の前壁は尿道と基部に隣接しています 膀胱、尿道の端部が下部に突き出ています。 前膣壁を膀胱から分離する結合組織の薄層は、膀胱膣中隔と呼ばれます。 正面から、膣は間接的に接続されています 後部恥骨嚢胞靭帯として知られている膀胱の基部にある筋膜結節による恥骨。 後ろでは、膣壁の下部が肛門管から会陰体によって隔てられています。 中央部は直腸に隣接し、上部は腹膜腔の直腸-子宮腔(ダグラス空間)に隣接しており、そこから腹膜の薄層によってのみ分離されています。
妊娠外の子宮(子宮)は、骨盤の正中線またはその近くにあります 膀胱前部と後部の直腸(図2.3を参照)。 子宮は、密な筋肉壁と三角形の内腔を備えた逆さの洋ナシの形をしており、矢状面では狭く、正面では広い。 子宮では、体、胃底、子宮頸部、峡部が区別されます。 膣アタッチメントラインは、子宮頸部を膣(膣)と膣上(膣上)のセグメントに分割します。 妊娠外では、湾曲した底は前方に向けられ、体は膣に対して鈍角を形成し(前方に傾く)、前方に曲がります。 子宮体の前面は平らで、膀胱の頂点に隣接しています。 後面は湾曲しており、上と後ろから直腸に面しています。
子宮頸部は下向きおよび後方に向けられており、膣の後壁と接触しています。 尿管は子宮頸部のすぐ横に比較的接近して接近します。 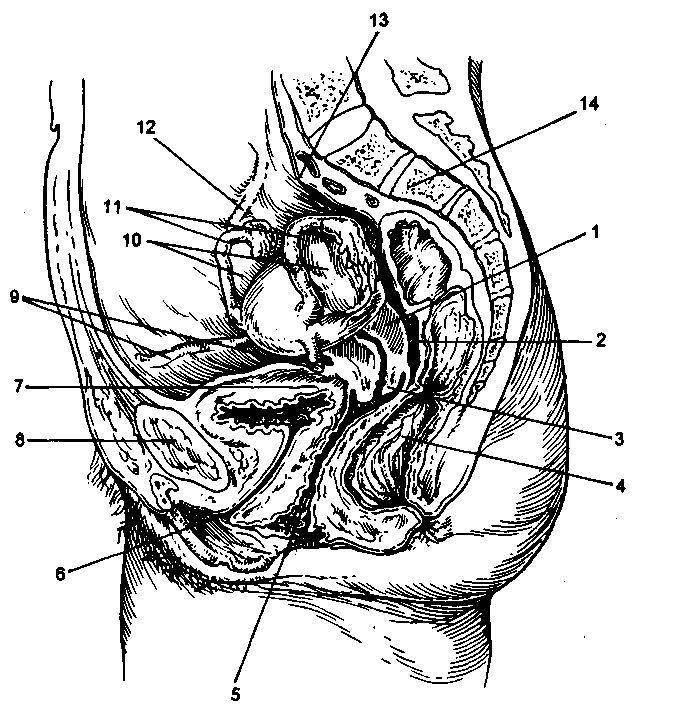
米。 2.3。 骨盤内臓器の比率(矢状カット)。
1-子宮; 2-直腸-子宮腔; 3-子宮頸部; 4-直腸; 5-膣; 6-尿道; 7-膀胱; 8-交感神経; 9-子宮の丸い靭帯; 10-卵巣; そして-卵管; 12-漏斗靭帯; 13-仙骨岬; 14-仙骨。
子宮の底を含む体は腹膜で覆われています。 正面では、峡部のレベルで、腹膜が曲がって膀胱の上面に通過し、浅い膀胱子宮腔を形成します。 背後では、腹膜は前方および上方に続き、峡部、子宮頸部の膣上部分、および膣の後円蓋を覆い、次に直腸の前面を通過して、深い直腸子宮腔を形成します。 子宮の体の長さは平均5cmです。子宮頸部と子宮頸部の全長は約2.5cm、直径は2cmです。体の長さと子宮頸部の比率は年齢と数によって異なります出生し、平均2:1です。
子宮の壁は、腹膜の薄い外層(漿膜(ペリメトリー)、平滑筋と結合組織の厚い中間層)、筋肉膜(子宮筋層)、および粘膜内膜(子宮内膜)で構成されています。 子宮の体には多くの筋線維が含まれており、子宮頸部に近づくにつれてその数は下向きに減少します。 首は同量の筋肉と結合組織で構成されています。 中腎傍管(ミュラー管)の合流部分からの発達の結果として、子宮壁の筋線維の位置は複雑です。 子宮筋層の外層には、主に上半身を横方向に走り、卵管の外側の縦筋層に接続する垂直繊維が含まれています。 中間層は子宮壁の大部分を含み、各チューブの内側の円形筋層に接続されているコイル状の筋線維のネットワークで構成されています。 支持靭帯の平滑筋線維の束が絡み合い、この層と融合します。 内層は、峡部および卵管の開口部で括約筋として機能することができる円形繊維で構成されています。
妊娠中の子宮腔は狭い隙間ですが、前壁と後壁は互いに密接に隣接しています。 空洞は逆三角形の形をしており、その底部は上部にあり、両側で卵管の開口部に接続されています。 頂点は子宮腔が子宮頸管に入る下にあります。 峡部の子宮頸管は圧迫されており、長さは6〜10mmです。 子宮頸管が子宮腔に入る場所は、内咽頭と呼ばれます。 子宮頸管は中央部でわずかに広がり、外部開口部で膣に通じています。
子宮の付属肢。 子宮付属器には卵管と卵巣が含まれ、一部の著者は子宮の靭帯装置についても言及しています。
卵管(子宮管)。 子宮体の両側には、横方向に長くて細い卵管(卵管)があります。 管は広間膜の上部を占め、卵巣の上で横方向に弧を描いて曲がり、次に卵巣の内側表面の後部の上を下って行きます。 管の内腔または管は、子宮腔の上隅から卵巣まで通過し、その経路に沿って横方向に直径が徐々に増加します。 妊娠以外では、伸ばされたチューブの長さは10cmです。4つのセクションがあります。 壁内サイト子宮壁の内側にあり、子宮腔に接続されています。 その内腔は最小の直径(1mm以下)を持っています。 子宮の外縁から横方向に伸びる狭い領域は、 地峡(地峡); さらに、パイプは膨張して曲がりくねり、形成されます アンプル、卵巣の近くで終わります じょうご。漏斗の周辺には、卵管の腹部開口部を囲む線毛があります。 1つまたは2つの線毛が卵巣と接触しています。 卵管の壁は3つの層で形成されています:主に腹膜(漿膜)、中間平滑筋層(筋サルピンクス)および粘膜(内サルピンクス)からなる外層。 粘膜は繊毛上皮で表され、縦方向のひだがあります。
卵巣(卵巣)。 女性の性腺は、楕円形またはアーモンド形の卵巣で表されます。 卵巣は卵管の湾曲した部分の内側に位置し、わずかに平らになっています。 平均して、それらの寸法は、幅2cm、長さ4cm、厚さ1cmです。卵巣は、通常、灰色がかったピンク色で、しわの寄った不均一な表面があります。 卵巣の縦軸はほぼ垂直で、上極点は卵管にあり、下極点は子宮に近い。 後部卵巣は自由であり、前部は腹膜の2層のひだ(卵巣の腸間膜(卵巣間膜))の助けを借りて子宮の広い靭帯に固定されています。 血管と神経がそれを通過し、卵巣門に到達します。 卵巣の上部極に取り付けられているのは、腹膜のひだです。これは、卵巣の血管と神経を含む卵巣を吊るす靭帯(ボロノタシス)です。 卵巣の下部は、線維筋性靭帯(卵巣自身の靭帯)によって子宮に付着しています。 これらの靭帯は、卵管が子宮体に近づく場所のすぐ下の角度で子宮の外側の端に接続します。
卵巣は胚性上皮で覆われており、その下には結合組織の層である卵巣の白膜があります。 卵巣では、外側の皮質層と内側の髄質層が区別されます。 血管と神経は延髄の結合組織を通過します。 皮質層では、結合組織の中で、多数の卵胞があります さまざまな段階発達。
女性の内臓の靭帯装置。子宮と卵巣の小さな骨盤、および膣と隣接する臓器の位置は、主に骨盤底の筋肉と筋膜の状態、および子宮の靭帯装置の状態に依存します(図2.2を参照)。 通常の位置では、卵管と卵巣を備えた子宮は、吊り下げ装置(靭帯)、固定装置(吊り下げられた子宮を固定する靭帯)、支持または支持装置(骨盤底)によって保持されます。
内性器の懸垂装置には、以下の靭帯が含まれています。
1.子宮の丸い靭帯(ligg。Teresuteri)。 それらは平滑筋と結合組織で構成され、長さ10〜12 cmの紐の形をしています。これらの靭帯は子宮の角から伸び、子宮の広い靭帯の前葉の下を通り、鼠径管の内部開口部に達します。 。 鼠径管を通過すると、子宮円索は恥骨と大陰唇の組織に扇状に分岐します。 子宮の丸い靭帯は、眼底を前方に引っ張ります(前方に傾いています)。
2.子宮の広い靭帯(ligg。Lataeuteri)。 これは、子宮の肋骨から骨盤の側壁に至る腹膜の重複です。 ワイドの上部に
子宮の靭帯は卵管を通過し、卵巣は裏側のシートにあり、シートの間には繊維、血管、神経があります。
3.卵巣の自身の靭帯(ligg。Ovariiproprii、s。Ligg。Suspensoriiovarii)は、卵管の排出場所の後ろと下の子宮底から始まり、卵巣に行きます。
4.卵巣を吊るす靭帯、または漏斗靭帯(ligg。Suspensoriumovarii、s.infundibulopelvicum)は、卵管から骨盤壁までの広い子宮靭帯の延長です。
子宮の固定装置は、子宮の下部から来る平滑筋線維の混合物を備えた結合組織コードです。
B)後方-直腸と仙骨(lig.sacrouterinum)まで。
仙骨靭帯は、体の移行領域の子宮の後面から子宮頸部まで伸び、両側の直腸を覆い、仙骨の前面に取り付けられています。 これらの靭帯は子宮頸部を後方に引っ張ります。
支持装置、または支持装置は、骨盤底の筋肉と筋膜で構成されています。 骨盤底は、内性器を正常な位置に保つために非常に重要です。 腹腔内圧が上昇すると、子宮頸部はサポートのように骨盤底に留まります。 骨盤底の筋肉は、性器と内臓が落ちるのを防ぎます。 骨盤底は、会陰の皮膚と粘膜、および筋膜の横隔膜によって形成されます。
会陰は、尿道、膣、肛門が位置する太ももと臀部の間のダイヤモンド形の領域です。 前部では、会陰は恥骨結合によって、後部では尾骨の端によって、外側坐骨結節によって囲まれています。 皮膚は会陰の外側と下を境界付け、下筋膜と上筋膜によって形成される骨盤横隔膜(骨盤筋膜)は上から会陰を深く境界付けます(図2.4)。
骨盤底は、2つの坐骨結節を結ぶ架空の線を使用して、解剖学的に2つの三角形の領域に分割されます。前部は泌尿生殖器領域、後部は肛門領域です。 会陰の中心、肛門と膣の開口部の間には、会陰の腱の中心と呼ばれる線維筋の形成があります。 この 腱センターいくつかの筋肉群と筋膜の付着部位です。
泌尿生殖器領域。泌尿生殖器領域では、坐骨と恥骨の下部枝の間に、「泌尿生殖器横隔膜」(diaphragma urogenitale)と呼ばれる筋膜の形成があります。 膣と尿道はこの横隔膜を通過します。 横隔膜は、外性器を固定するための基礎として機能します。 下から、尿生殖横隔膜は、尿生殖領域を重要な2つの高密度の解剖学的層に分割する尿生殖横隔膜の下部筋膜を形成する白っぽいコラーゲン繊維の表面によって制限されます 臨床的な意義、-表面的で深いセクション、または股ポケット。
浅会陰隙。表層部は、泌尿生殖器横隔膜の下部筋膜の上に位置し、両側に膣の前庭の大きな腺、上に骨格筋が横たわっている陰核脚、前庭球の球根が含まれています-上に横たわる海綿状(前庭球-海綿体)の筋肉、および小さな表在性の横筋。 坐骨海綿体筋は陰核の脚を覆い、脚を坐骨神経枝に押し付けて勃起組織からの血液の流出を遅らせるため、その勃起を維持する上で重要な役割を果たします。 球根状の骨格筋は腱から始まります- 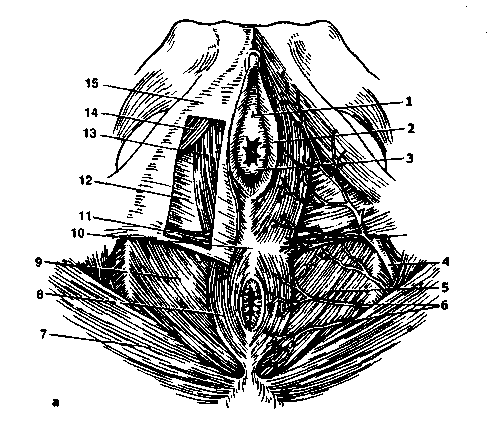
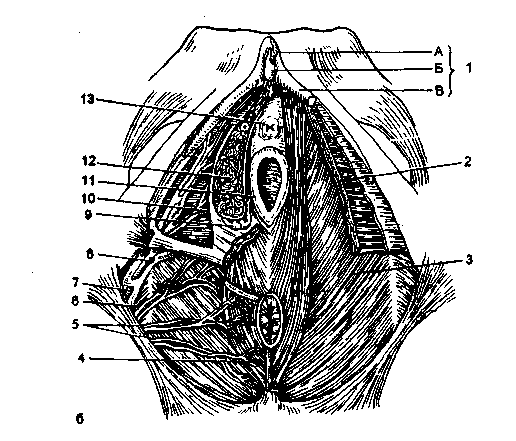
米。 2.4。 女性の股間。
A-泌尿生殖器横隔膜の表層部:1-尿道の外部開口部、2-小陰唇、3-会陰、4-内生殖器動脈、5-肛門を持ち上げる筋肉、6-下痔核動脈、7-会陰部筋肉、8-肛門の外筋膜、9-骨盤底筋膜の下部筋膜、10-会陰の腱中心、11-会陰の外部横筋膜、12-泌尿生殖器横隔膜の下部筋膜、13-球根状スポンジ筋肉、14-ischiocavernous筋肉、15-会陰の表在性筋膜; b *-泌尿生殖器横隔膜の深部:1-クリトリス:A-体、B-頭、B-脚; 2-尿生殖器横隔膜、3-骨盤横隔膜、4-肛門の外括約筋の筋肉、5-下痔核動脈、6-内閉鎖筋、7-内閉鎖筋、8-会陰動脈、9-大きな前庭腺、 10-前庭動脈、11-膣壁、12-前庭の球根、13-尿道。
会陰の中心と肛門の外括約筋は、後ろから膣の下部の周りを通り、前庭球を覆い、会陰体に入ります。 筋肉は、膣の下部を圧迫する括約筋として機能することができます。 薄い板のように見える会陰の発達が不十分な表在性横筋は、坐骨結節近くの坐骨の内面から始まり、横方向に進み、会陰体に入る。 表層領域のすべての筋肉は会陰の深い筋膜で覆われています。
会陰の深い部分。会陰の深部は、泌尿生殖器横隔膜の下部筋膜と尿生殖器横隔膜の不明瞭な上部筋膜の間にあります。 泌尿生殖器の横隔膜は2層の筋肉で構成されています。 泌尿生殖器横隔膜の筋線維は主に横方向に位置し、両側の坐骨恥骨枝から離れて正中線に沿って接続されています。 尿生殖器横隔膜のこの部分は、会陰の深部横筋と呼ばれます(m。Transversusperineiprofundus)。 尿道括約筋の繊維の一部は尿道の上に弧を描いて上昇し、他の部分はその周りに円形に配置され、尿道の外括約筋を形成します。 尿道括約筋の筋線維も膣の周りを通過し、尿道の外部開口部が位置する場所に集中します。 筋肉は、膀胱がいっぱいのときに排尿のプロセスを抑制するのに重要な役割を果たし、尿道の任意の収縮剤です。 深会陰横筋は、膣の後ろの会陰体に入ります。 したがって、両側の収縮により、この筋肉は会陰とそれを通過する内臓構造を支えます。
泌尿生殖器横隔膜の前縁に沿って、その2つの筋膜が合流し、横会陰靭帯を形成します。 この筋膜肥厚の前には、恥骨結合の下端に沿って走る弓状の恥骨靭帯があります。
肛門(肛門)エリア。肛門(肛門)領域には、肛門、肛門の外括約筋、および坐骨直腸窩が含まれます。 肛門は会陰の表面にあります。 肛門の皮膚は色素沈着しており、皮脂腺と汗腺が含まれています。 肛門の括約筋は、骨格筋線維の表層部と深部で構成されています。 皮下部分は最も表面的で、直腸の下壁を囲み、深部は肛門挙筋と合流する円形の繊維で構成されています。 括約筋の表層部は、主に肛門管に沿って走り、肛門の前と後ろで直角に交差し、会陰の前と後ろに入る筋線維で構成されています。肛門-尾骨体、または肛門-尾骨靭帯。 肛門の外側は縦方向のスリット開口部であり、これはおそらく肛門の外括約筋の多くの筋線維の前後方向によって説明されます。
直腸直腸窩は、脂肪で満たされたくさび形の空間であり、外側から皮膚で囲まれています。 革はくさびのベースを形成します。 窩の垂直側壁は、内閉鎖筋によって形成されます。 傾斜した肩甲挙筋には肛門挙筋が含まれています。 坐骨神経痛組織は、排便時に直腸と肛門管を拡張させることができます。 窩とそれに含まれる脂肪組織は、泌尿生殖器の横隔膜まで前方に深く位置していますが、肛門を持ち上げる筋肉の下にあります。 この領域はフロントポケットと呼ばれます。 背後では、窩の脂肪組織が仙骨靭帯の領域の大殿筋まで深く伸びています。 横方向では、窩は内閉鎖筋の下部を覆う坐骨と閉鎖筋膜によって囲まれています。
血液供給、リンパ排液および生殖器の神経支配。 血液供給(図2.5、2.6)外性器の主に内性器(陰部)動脈によって実行され、部分的にのみ枝によって実行されます 大腿動脈.
内性器動脈(a.pudenda interna)は 主動脈会陰。 これは、内腸骨動脈(a.iliaca interna)の枝の1つを表します。 骨盤腔を出て、大坐骨孔の下部を通過し、次に坐骨脊椎の周りを曲がり、坐骨直腸窩の側壁に沿って進み、小坐骨孔を横方向に横切る。 その最初の枝は下直腸動脈(下直腸動脈)です。 坐骨神経痛を通過して、肛門周辺の皮膚や筋肉に血液を供給します。 会陰枝は浅会陰隙の構造を供給し、大陰唇と小陰唇に向かう後部会陰の形で続きます。 深会陰隙に入る内性器動脈は、いくつかの断片に分岐し、前庭球に血液、前庭の大きな腺、および尿道を供給します。 終わりに、それは陰核の深くて背側の動脈に分かれて、恥骨結合の近くでそれに近づきます。
外部(表在性)生殖器動脈(r.pudenda externa、s.superficialis)は、大腿動脈(a.femoralis)の内側から離れ、大陰唇の前部に血液を供給します。 外部(深部)生殖器動脈(r.pudenda externa、s.profunda)も大腿動脈から離れていますが、より深く、より遠位にあります。 太ももの内側の広い筋膜を通過した後、大陰唇の外側部分に入ります。 その枝は前陰唇動脈と後陰唇動脈に入ります。
会陰を通過する静脈は、主に内腸骨静脈の枝です。 ほとんどの場合、それらは動脈に付随します。 例外は陰核の深い背側静脈で、陰核の勃起組織から恥骨結合の下の隙間を通って膀胱頸部の周りの静脈叢に血液を排出します。 外性器静脈は大陰唇から血液を排出し、横方向に通過して脚の大伏在静脈に入ります。
内性器への血液供給は、主に大動脈(総腸骨動脈と内腸骨動脈のシステム)から行われます。
子宮への主な血液供給は、内腸骨(下腹部)動脈(a.iliacainterna)から離れる子宮動脈(a.uterina)によって提供されます。 症例の約半分では、子宮動脈は内腸骨動脈から自発的に離れますが、臍帯、内性器、表在性嚢胞動脈から始まることもあります。
子宮動脈は骨盤外側壁まで下がってから、尿管の上にある前方および内側を通過し、そこに独立した枝を与えることができます。 広い子宮靭帯の基部で、それは子宮頸部に向かって内側に曲がります。 子宮傍組織では、動脈は付随する静脈、神経、尿管、基靭帯に接続しています。 子宮動脈は子宮頸部に近づき、いくつかの曲がりくねった貫通枝を通して子宮頸部に供給します。 次に、子宮動脈は、1つの大きな、非常に複雑な上行枝と1つ以上の小さな下行枝に分かれて、上部膣と隣接する膀胱に供給します。 主な上行枝は子宮の外側の端に沿って上向きに走り、その体に向かって弧状の枝を送ります。 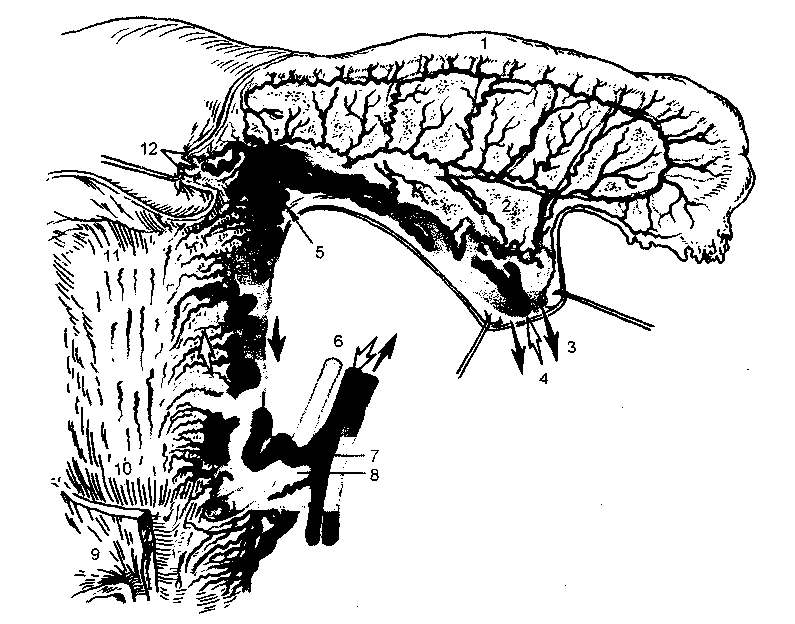
米。 2.5。 性器への血液供給。
1-卵管; 2-卵巣; 3-卵巣静脈; 4-卵巣動脈; 5-子宮および卵巣血管の吻合; 6-尿管; 7-子宮動脈; 8-子宮静脈; 9-膀胱の壁; 10-子宮頸部; 11-子宮の体; 12-子宮円索。
これらの弓状動脈は漿膜の下の子宮を囲んでいます。 一定の間隔で、放射状の枝がそれらから離れ、子宮筋層の絡み合っている筋線維に浸透します。 出産後、筋線維は収縮し、結紮糸として作用して、橈骨枝を圧迫します。 弓状動脈のサイズは正中線に向かって急速に減少するため、正中線の子宮切開では側方切開よりも出血が少なくなります。 子宮動脈の上行枝は卵管に近づき、上部で横方向に回転し、卵管枝と卵巣枝に分かれます。 卵管枝は、卵管(mesosalpinx)の腸間膜を横方向に走っています。 卵巣枝は卵巣の腸間膜(卵巣間膜)に向けられており、大動脈から直接伸びる卵巣動脈と吻合します。
卵巣は、腹部大動脈から左に伸びる卵巣動脈(a.ovarica)から供給され、腎動脈(a.renalis)から供給されることもあります。 尿管と一緒に下がると、卵巣動脈は、卵巣を広い子宮靭帯の上部に吊るす靭帯に沿って通過し、卵巣と管のための枝を与えます。 卵巣動脈の末端部分は、子宮動脈の末端部分と吻合します。
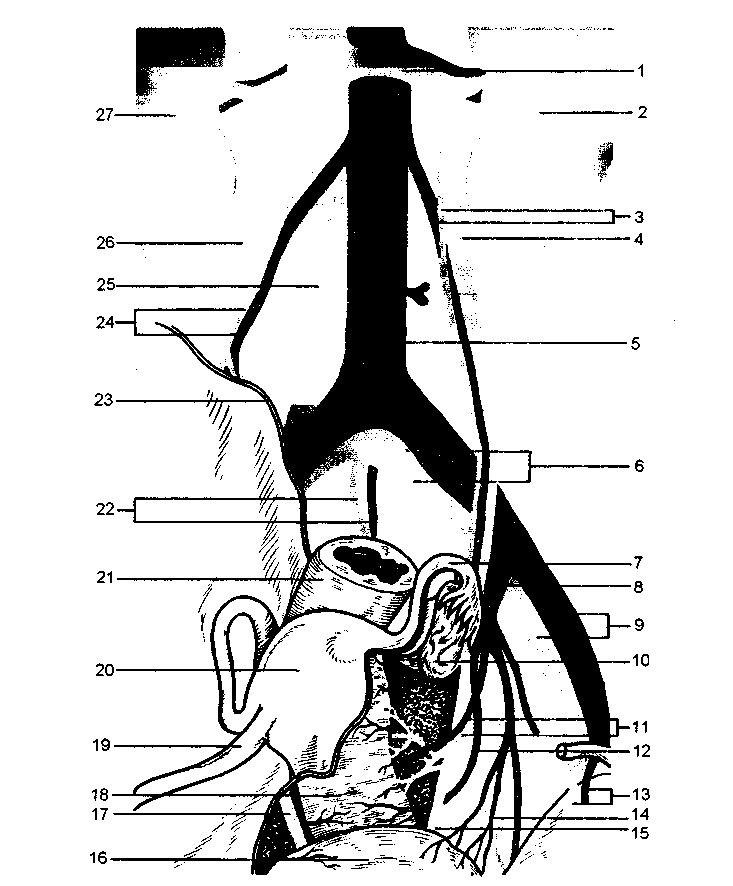
米。 2.6。 小骨盤の内性器、尿管、血管の関係。
1-左腎静脈; 2-左腎臓; 3-左卵巣の静脈と動脈; 4-左尿管; 5-大動脈の腹部; 6-総腸骨動脈および静脈; 7-卵管; 8-内腸骨動脈; 9-外腸骨動脈および静脈; 10-左卵巣; 11-子宮動脈および子宮静脈; 12-下部嚢胞動脈(膣枝); 13-下腹壁動脈および静脈; 14-上尿路; 15-左尿管; 16-膀胱; 17-右尿管; 18-膣; 19-子宮円索; 20-子宮の体; 21-直腸; 22-正中仙骨静脈および動脈; 23-壁側腹膜の端(断面); 24-右卵巣の動脈と静脈; 25-下大静脈; 26-右尿管; 27-右腎臓。
子宮と生殖器の動脈に加えて、下直腸動脈と中直腸動脈の枝も膣への血液供給に関与しています。 生殖器の動脈は、対応する静脈を伴います。
生殖器の静脈系は非常に強く発達しています;全長 静脈血管...互いに広く吻合された静脈叢が存在するため、動脈の長さを大幅に超えています。 静脈叢は、陰核、前庭球の端、膀胱の周り、子宮と卵巣の間にあります。
リンパ系生殖器は、曲がりくねったリンパ管、神経叢、および多くのリンパ節の密なネットワークで構成されています。 リンパ管とリンパ節は主に 血管.
外性器と膣の下3分の1からリンパを排出するリンパ管は鼠径リンパ節に行きます。 膣と子宮頸部の中央上部3分の1から伸びるリンパ経路は、下腹部と腸骨の血管に沿って位置するリンパ節に行きます。
壁内神経叢は、子宮内膜および子宮筋層から漿液性神経叢にリンパ液を運び、そこからリンパ液が遠心性血管を通って流れます。 子宮の下部からのリンパ液は、主に仙骨、外腸骨動脈、および総腸骨リンパ節に入ります。 一部のリンパ液は、腹部大動脈に沿った腰椎下部リンパ節および表在性鼠径リンパ節にも流れます。 子宮の上部からのリンパ液の大部分は、子宮の広い靭帯に横方向に流れ込み、そこで卵管と卵巣から集められたリンパ液とつながります。 さらに、卵巣を吊るす靭帯を通って、卵巣血管に沿って、リンパは下腹部大動脈に沿ってリンパ節に入ります。 リンパ液は卵巣から卵巣動脈に沿った血管を通って排出され、大動脈と下大静脈にあるリンパ節に行きます。 これらのリンパ叢の間には関連性があります-リンパ吻合。
神経支配中女性の生殖器には、自律神経系の交感神経と副交感神経の部分、および脊髄神経が関与しています。
自律神経系の交感神経部分の線維は、生殖器を神経支配し、大動脈および腹腔(「太陽」)神経叢に由来し、下降して、V腰椎のレベルで上部下腹部神経叢(上神経叢)を形成します。 。 左右の下腹部神経叢(骨盤神経叢の不吉な神経叢と下のデクスター神経叢)を形成する線維は、そこから離れます。 これらの神経叢からの神経線維は、強力な子宮膣神経叢、または骨盤神経叢(神経叢、骨盤神経叢)に行きます。
子宮膣神経叢は、咽頭内および子宮頸管のレベルで、子宮の外側および子宮の後ろのパラメトリック組織に位置しています。 この神経叢には、自律神経系の副交感神経部分に属する骨盤神経(n.pelvicus)の枝が接近します。 子宮膣神経叢から伸びる交感神経および副交感神経線維は、膣、子宮、卵管の内部部分、および膀胱を神経支配します。
卵巣は、卵巣神経叢(神経叢)からの交感神経と副交感神経によって神経支配されています。
外性器と骨盤底は主に陰部神経(n.pudendus)によって神経支配されています。
骨盤組織。骨盤内臓器の血管、神経、リンパ経路は、腹膜と骨盤底筋膜の間にある組織を通過します。 繊維はすべての骨盤内臓を取り囲んでいます。 一部の領域では緩んでおり、他の領域では繊維状のストランドの形をしています。 組織の次のスペースがあります:眼周囲、小胞前および小胞周囲、腸周囲、膣。 骨盤組織は内性器を支え、そのすべてのセクションが相互接続されています。
2.1.3。 産科の観点からの骨盤
大きな骨盤は子供の誕生に不可欠ではありません。 生まれた胎児の障害となる産道の骨の基礎は、小さな骨盤です。 しかし、大きな骨盤の大きさから、小さな骨盤の形や大きさを間接的に判断することができます。 大小の骨盤の内面には筋肉が並んでいます。
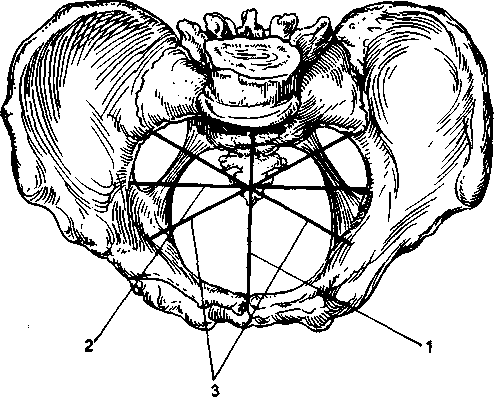
小さな骨盤腔骨盤の入口と出口の平面によって制限された、骨盤の壁の間に囲まれた空間と呼ばれます。 円柱の形をしており、前から後ろに切り詰められており、胸に面する前部は、仙骨に面する後部のほぼ3分の1になっています。 骨盤腔のこの形状に関連して、そのさまざまなセクションは異なる形状とサイズを持っています。 これらの分割は、小さな骨盤の内面の識別ポイントを通過する仮想平面です。 小さな骨盤では、次の平面が区別されます:入口の平面、広い部分の平面、狭い部分の平面、および出口の平面(表2.1;図2.7)。
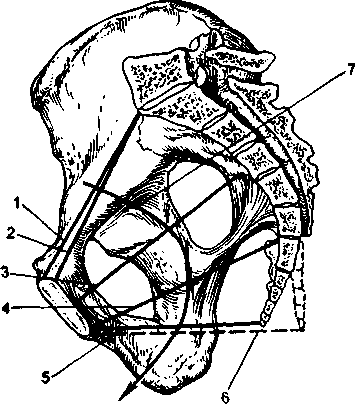 |  |
| 米。 2.7。 女性の骨盤(矢状カット)。 1-解剖学的コンジュゲート; 2-真の共役; 3-骨盤腔の広い部分の平面の直線サイズ。 4-骨盤腔の狭い部分の平面の直接サイズ。 5-尾骨の通常の位置にある小さな骨盤の出口の直線サイズ。 6-尾骨が後方に曲がっている小さな骨盤の出口の直接のサイズ。 7-骨盤のワイヤー軸。 | 米。 2.8。 骨盤への入り口の平面の寸法。 1-ストレートサイズ(真の共役); 2-横方向の寸法; 3-斜めの寸法。 |
エントリープレーン小さな骨盤に入ると、恥骨弓の上部内側の端、名前のない線、および岬の上部を通過します。 入口面では、以下の寸法が区別されます(図2.8)。
ストレートサイズ-恥骨弓の上部内側エッジの中央と岬の最も目立つポイントの間の最短距離。 この距離はアオミドロと呼ばれます。 それは11cmに等しいです。また、区別して 解剖学的コンジュゲート-上部の中央からの距離 エッジ岬の同じポイントへの陰部弧; 真の共役より0.2-0.3cm長い(図2.7を参照)。
横方向の寸法-反対側の匿名の線の最も遠い点の間の距離。 それは13.5cmです。この寸法は、岬に近い、真の共役を偏心して直角に交差します。
斜めの寸法-左右。 右の斜めのサイズは、それぞれ、右の仙腸関節から左の腸骨-恥骨結節になり、左の斜めのサイズは、左の仙腸関節から右の腸骨-恥骨結節になります。 これらの寸法はそれぞれ12cmです。
与えられた寸法からわかるように、入口面は横楕円形をしています。
骨盤腔の広い部分の平面は、恥骨弓の内面の中央を横から、寛骨臼(椎弓板)の窩の下にある滑らかなプレートの中央を通り、後ろを通ります。 -IIとIIIの仙椎の間の関節を介して。
広い部分の平面では、次の寸法が区別されます。
ストレートサイズ-恥骨弓の内面の中央からIIとIIIの仙椎の間の関節まで。 12.5cmです。
両側の寛骨臼プレートの最も遠い点を結ぶ横方向の寸法は12.5cmです。
広い部分の平面は円に近い形をしています。
狭い部分の平面骨盤腔は、前部から陰部関節の下端を通り、側面から-坐骨棘を通り、後ろから-仙尾骨関節を通ります。
狭い部分の平面では、次の寸法が区別されます。
ストレートサイズ-恥骨結合の下端から仙尾骨関節まで。 11cmです。
横方向の寸法は、坐骨棘の内面の間にあります。 それは10.5cmに等しいです。
出口面小さな骨盤は、小さな骨盤の他の平面とは異なり、坐骨結節を結ぶ線に沿ってある角度で収束する2つの平面で構成されます。 それは前部を恥骨弓の下端を通り、側面を-坐骨結節の内面を通り、そして後ろを-尾骨の頂点を通ります。
次の寸法は、出口面で区別されます。
ストレートサイズ-陰部関節の下端の中央から尾骨の頂点まで。 9.5cmに相当します(図2.9)。 尾骨のある程度の可動性のために、出口のまっすぐなサイズは、胎児の頭の通過中に出産時に1〜2cm長くなり、11.5cmに達する可能性があります(図2.7を参照)。
横方向の寸法は、坐骨結節の内面の最も遠い点の間です。 11cmです(図2.10)。
表2.1。 小さな骨盤の平面と寸法
| 骨盤の平面 | 寸法、cm |
|||||
| 真っ直ぐ | 横 | 斜め |
||||
| 盆地への入り口 | 11 | 13-13,5 | 12-12,5 |
|||
| 骨盤腔の最も広い部分 | 12,5 | 12,5 | 13(条件付き) |
|||
| 骨盤腔の狭い部分 | 11-11,5 | 10,5 | - |
|||
| 骨盤の出口 | 9,5-11,5 | 11 | - |
|||
|
|
| |||||
| 米。 2.9。骨盤出口の直接サイズ(測定)。 | 米。 2.10。骨盤出口の横方向の寸法の測定。 | |||||
ロシアの産科の創設者、特にA.Ya. Krassovskyが参加したこの古典的な飛行機のシステムでは、産道に沿って胎児の提示部分の前進を正しくナビゲートし、それを回転させることができますこの間に作ります。
V 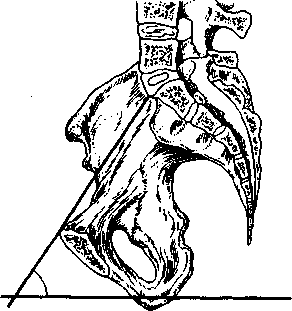
米。 2.11。 骨盤の傾斜角
小さな骨盤の平面のすべての直線寸法は、陰部の関節の領域で収束しますが、仙骨の領域で発散します。 骨盤面のすべての直線寸法の中点を結ぶ線は円弧であり、前面が凹状で背面が湾曲しています。 この線は有線骨盤軸と呼ばれます。 胎児が産道を通過するのはこの線に沿って行われます(図2.7を参照)。
女性が立っているときの骨盤の傾斜角度(入口の平面と地平線の平面の交点(図2.11))は、体格によって異なり、45〜55°の範囲です。 仰向けになっている女性に腰を強く引き寄せて胸を持ち上げるように頼むと減少する可能性があり、逆に、ローラーのような硬い枕を腰の下に置くと増加する可能性があります。これは、胸の下方への逸脱につながります。 女性が半座位またはしゃがむ姿勢をとると、骨盤の傾斜角の減少も達成されます
2.2。 女性の生殖システムの生理学。 月経周期
女性の生殖機能は、主に卵巣と子宮の活動によって実行されます。卵巣は卵巣で成熟するため、子宮では、卵巣から分泌されるホルモンの影響下で、卵巣の知覚に備えて変化が起こります。受精した卵巣。 生殖期間は、女性の体が子孫を生殖する能力によって特徴付けられます。 この期間の期間は17-18から45-50年です。 生殖または出産の期間の前に、女性の人生の次の段階があります。子宮内。 新生児(1歳まで); 小児期(8-10歳まで); 思春期前および思春期の年齢(17〜18歳まで)。 生殖期は閉経に変わり、閉経前、閉経、閉経後が区別されます。
月経周期-女性の体の複雑な生物学的プロセスの症状の1つ。 月経周期は、すべてのリンクの周期的な変化によって特徴付けられます 生殖システム、その外部症状は月経です。
月経-これは女性の生殖管からの血の分泌物であり、二相性月経周期の終わりに子宮内膜の機能層が拒絶された結果として定期的に発生します。 最初の月経(初潮)は10〜12歳で発生しますが、その後1〜1。5年は月経が不規則になることがあり、その後、定期的な月経周期が確立されます。
月経の初日は、通常、月経周期の初日と見なされます。 したがって、サイクルの期間は、次の2つの月経の最初の日の間の時間です。 女性の60% 平均デュレーション月経周期は28日で、21日から35日まで変動します。 月経日の失血量は40〜60ml、平均50mlです。 通常の月経の期間は2〜7日です。
卵巣。月経周期の間に、卵胞は卵巣で成長し、卵子は成熟し、その結果、受精の準備が整います。 同時に、性ホルモンは卵巣で生成され、受精卵を受け取ることができる子宮の内壁に変化をもたらします。
性ホルモン(エストロゲン、プロゲステロン、アンドロゲン)はステロイドであり、卵胞の顆粒膜細胞、内層と外層の細胞がそれらの形成に関与しています。 卵巣によって合成される性ホルモンは、標的の組織や臓器に影響を与えます。 これらには、生殖器、主に子宮、乳腺、海綿状骨、脳、内皮および血管平滑筋細胞、心筋、皮膚およびその付属物(毛包および皮脂腺)などが含まれます。標的へのホルモンの直接接触および特異的結合細胞は、対応する受容体との相互作用の結果です。
生物学的効果は、エストラジオールとテストステロンの遊離(非結合)画分(1%)によって提供されます。 卵巣ホルモンの大部分(99%)が結合しています。 輸送は、特別なタンパク質(ステロイド結合グロブリンおよび非特異的輸送システム)、アルブミンおよび赤血球によって実行されます。 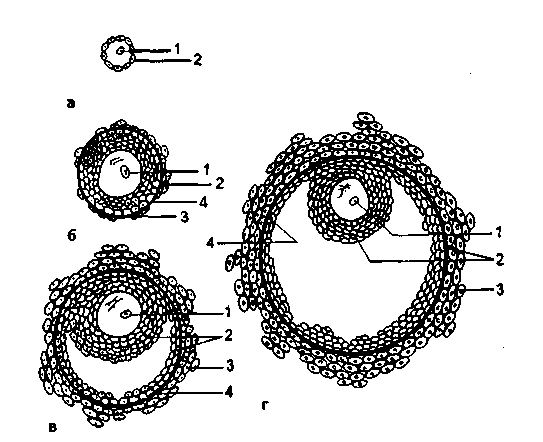
米。 2.12。優勢な濾胞の発達段階。
A-原始濾胞; b-前胞状卵胞; c-胞状卵胞; d-排卵前卵胞:1-卵母細胞、2-顆粒膜細胞(顆粒帯)、3-卵胞膜、4-基底膜。
エストロゲンホルモンは、性器の形成、思春期の第二次性徴の発症に寄与します。 アンドロゲンは恥骨と脇の下の髪の外観に影響を与えます。 プロゲステロンは月経周期の分泌期を制御し、着床のために子宮内膜を準備します。 性ホルモンは妊娠と出産の発達に重要な役割を果たします。
卵巣の周期的な変化には、3つの主要なプロセスが含まれます。
1.卵胞の成長と優勢な卵胞の形成。
2.排卵。
3.黄体の形成、発達および退行。
少女の誕生時には、卵巣に200万個の卵胞があり、その99%が生涯にわたって閉鎖状態にあります。 閉鎖のプロセスは、その発達の段階の1つで卵胞の逆発達として理解されています。 初潮の時までに、卵巣には約20万から40万の卵胞が含まれ、そのうち300から400は排卵の段階まで成熟します。
卵胞発達の次の主要な段階を区別するのが通例です(図2.12):原始卵胞、前胞状卵胞、胞状卵胞、排卵前卵胞。
原始卵胞は、卵胞および顆粒(顆粒)上皮に位置する未成熟卵細胞で構成されています。 外では、卵胞は結合膜(卵胞膜)に囲まれています。 各月経周期の間に、3〜30個の原始卵胞が成長し始め、前胞状または原発卵胞を形成します。
前胞状卵胞。 成長が始まると、原始卵胞は前胞状段階に進み、卵母細胞は成長し、透明帯と呼ばれる膜に囲まれます。 顆粒膜上皮の細胞は増殖し、周囲の間質から卵胞膜が形成されます。 この成長は、エストロゲン産生の増加を特徴としています。 前胞状卵胞の顆粒膜層の細胞は、3つのクラスのステロイドを合成することができますが、エストロゲンはアンドロゲンやプロゲステロンよりもはるかに多く合成されます。
アントラル、またはセカンダリ、follおよびkul。 それはさらなる成長によって特徴づけられます:卵胞液を生成する顆粒膜層の細胞の数が増加します。 卵胞液は顆粒膜層の細胞間空間に蓄積し、空洞を形成します。 卵胞形成のこの期間(月経周期の8-9日目)の間に、性ステロイドホルモン、エストロゲンおよびアンドロゲンの合成が注目されます。
性ホルモンの合成に関する現代の理論によれば、アンドロゲンは卵胞膜細胞で合成されます-アンドロステンジオンとテストステロン。 次に、アンドロゲンは顆粒膜層の細胞に入り、それらの中でそれらはエストロゲンに芳香化されます。
優勢な濾胞..。 原則として、そのような卵胞の1つは、多くの胞状卵胞から形成されます(サイクルの8日目までに)。 それは最も大きく、顆粒膜層の細胞とFSH、LHの受容体の最大数を含んでいます。 優勢な卵胞は、血管が豊富な卵胞膜を持っています。 卵巣における優勢な排卵前卵胞の成長と発達とともに、残りの(90%)成長中の卵胞の閉鎖のプロセスが並行して起こります。
月経周期の最初の日の優勢な濾胞は2mmの直径を持っています、そしてそれは排卵の時までに14日以内に平均21mmに増加します。 この間、卵胞液の量が100倍に増加します。 それはエストラジオールとFSHの含有量を急激に増加させ、成長因子も決定されます。
排卵は、卵胞前の優性(三次)卵胞の破裂とそこからの卵子の放出です。 排卵の時までに、減数分裂のプロセスは卵母細胞で起こります。 排卵は、卵胞膜細胞を取り巻く破壊された毛細血管からの出血を伴います。 排卵は、エストラジオールの排卵前のピークが形成されてから24〜36時間後に発生すると考えられています。 コラゲナーゼ酵素の影響下で、排卵前の卵胞壁の菲薄化と破裂が起こります。 卵胞液に含まれるプロスタグランジンF2aおよびE2によっても特定の役割が果たされます。 顆粒膜細胞で産生されるタンパク質分解酵素; オキシトシンとリラキシン。
卵子の放出後、結果として生じる毛細血管はすぐに卵胞腔に成長します。 顆粒膜細胞は黄体形成を起こします:細胞質の体積がそれらの中で増加し、脂質封入体が形成されます。 LHは、顆粒膜細胞のタンパク質受容体と相互作用し、黄体形成のプロセスを刺激します。 このプロセスは黄体の形成につながります。
黄体は、月経周期の長さに関係なく、14日間機能する一過性の内分泌腺です。 妊娠がない場合、黄体は退行します。
米。 2.13。月経周期中の血漿中のホルモンの含有量。
したがって、卵巣は主な女性の性ステロイドホルモンであるエストラジオールとプロゲステロン、およびアンドロゲンを合成します。
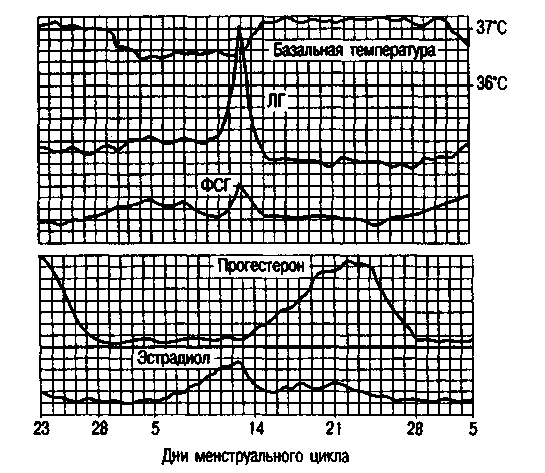
月経の初日から排卵の瞬間まで続く月経周期のフェーズIでは、体はエストロゲンの影響下にあり、フェーズII(排卵から月経の開始まで)では、細胞によって分泌されるプロゲステロン黄体のはエストロゲンに参加します。 月経周期の最初の段階は卵胞期または卵胞期とも呼ばれ、周期の2番目の段階は黄体期です。
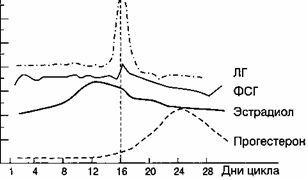
月経周期の間、末梢血中のエストラジオールの含有量には2つのピークがあります。1つ目は顕著な排卵前周期であり、2つ目はそれほど顕著ではなく、月経周期の第2段階の中間です。 サイクルの第2フェーズで排卵した後、プロゲステロンが主なものであり、その最大量は排卵後4〜7日目に合成されます(図2.13)。
卵巣でのホルモンの周期的な分泌は、子宮の内層の変化を決定します。
子宮内膜(子宮内膜)の周期的な変化。子宮内膜は次の層で構成されています。
1.月経中に拒絶されない基底層。 月経周期中の細胞から、子宮内膜の層が形成されます。
2.子宮腔の内側を覆うコンパクトな上皮細胞からなる表層。
3.中間層または海綿状の層。
最後の2つの層は機能層を構成します。機能層は月経周期の間に大きな周期的変化を受け、月経中に拒絶されます。
月経周期のフェーズIでは、子宮内膜は腺と間質の薄い層です。 サイクル中の子宮内膜の変化には、次の主な段階があります。
1)増殖の段階;
2)分泌の段階;
3)月経。
増殖期。 成長中の卵胞によるエストラジオールの分泌が増加すると、子宮内膜は増殖性の変化を起こします。 基底層の細胞は活発に増殖します。 細長い管状腺を備えた新しい表面の緩い層が形成されます。 この層はすぐに4〜5倍厚くなります。 円柱上皮で裏打ちされた管状腺は細長くなっています。
分泌段階。 卵巣周期の黄体期では、プロゲステロンの影響下で、腺の屈曲度が増加し、その内腔が徐々に拡大します。 間質の細胞は、体積が増加し、互いに接近します。 腺の分泌が増加します。 腺の内腔には大量の分泌物が見られます。 分泌の強さに応じて、腺は強く複雑なままであるか、鋸歯状になります。 間質の血管新生が増加しています。 分泌の初期、中期、後期を区別します。
月経。 これは、子宮内膜の機能層の拒絶反応です。 月経の開始とプロセスの根底にある微妙なメカニズムは不明です。 月経の開始の内分泌基盤は、黄体の退行によるプロゲステロンおよびエストラジオールレベルの顕著な減少であることが見出された。
月経に関与する主な局所メカニズムは次のとおりです。
1)らせん状動脈の色調の変化;
2)子宮内の止血のメカニズムの変化;
3)子宮内膜細胞のリソソーム機能の変化;
4)子宮内膜の再生。
月経の開始に先立って、らせん状細動脈が激しく狭くなり、子宮内膜の虚血および落屑につながることが確立されている。
月経周期の間に、子宮内膜細胞のリソソームの内容は変化します。 リソソームには酵素が含まれており、その一部はプロスタグランジンの合成に関与しています。 プロゲステロンレベルの低下に応じて、これらの酵素の放出が増加します。
子宮内膜の再生は、月経の最初から観察されます。 月経の24時間目の終わりまでに、子宮内膜の機能層の2/3が拒絶されます。 基底層には、子宮内膜再生の基礎となる間質上皮細胞が含まれています。これは通常、サイクルの5日目までに完全に完了します。 並行して、血管新生は、破裂した細動脈、静脈、および毛細血管の完全性の回復とともに完了します。
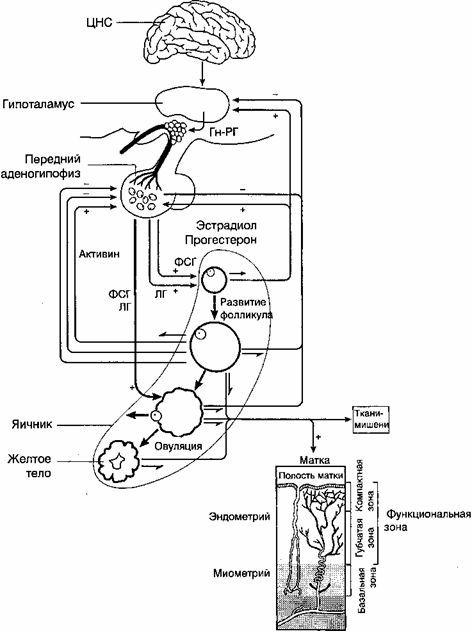
卵巣と子宮の変化は、大脳皮質、視床下部、下垂体などの月経機能を調節するシステムの二相性活動の影響下で発生します。 したがって、女性の生殖器系には、大脳皮質、視床下部、下垂体、卵巣、子宮の5つの主要な関連性があります(図2.14)。 生殖器系のすべてのリンクの相互関係は、性ホルモンと性腺刺激ホルモンの両方の受容体の存在によって保証されています。
中枢神経系の役割について生殖器系の機能の調節においては、長い間知られていました。 これは、さまざまな急性および慢性のストレス時の排卵の違反、気候帯を変更するときの月経周期の違反、仕事のリズムによって証明されました。 戦時中の月経の停止はよく知られています;子供を産むことを熱望している精神的に不安定な女性も月経をやめるかもしれません。
大脳皮質および視床下部外の大脳構造(大脳辺縁系、海馬、扁桃腺など)では、エストロゲン、プロゲステロン、アンドロゲンの特定の受容体が同定されています。 これらの構造では、神経ペプチド、神経伝達物質、およびそれらの受容体の合成、分泌、代謝が起こり、視床下部放出ホルモンの合成と分泌に選択的に影響を及ぼします。
性ステロイドと組み合わせて、それらは機能します 神経伝達物質:ノルエピネフリン、ドーパミン、ガンマアミノ酪酸、アセチルコリン、セロトニン、メラトニン。 ノルエピネフリンは、視床下部前部のニューロンからのゴナドトロピン放出ホルモン(GTRH)の放出を刺激します。 ドーパミンとセロトニンは、月経周期のさまざまな段階でGTRH産生の頻度と振幅を減少させます。
ニューロペプチド(内因性オピオイドペプチド、ニューロペプチドY、コルチコトロピン放出因子およびガラニン)も生殖器系の機能に影響を及ぼし、したがって視床下部の機能に影響を及ぼします。 3種類の内因性オピオイドペプチド(エンドルフィン、エンケファリン、ダイノルフィン)は、脳内のアヘン剤受容体に結合することができます。 内因性オピオイドペプチド(EOP)は、フィードバックメカニズムによってGTRH含有量に対する性ホルモンの影響を調節し、視床下部でのGTRHの分泌をブロックすることにより、下垂体、特にLHによる性腺刺激ホルモンの分泌をブロックします。
神経伝達物質と神経ペプチドの相互作用は、視床下部によるGTRHの合成と分泌に影響を与える、生殖年齢の女性の体内で定期的な排卵周期を提供します。
視床下部には、刺激(リベリン)および遮断(スタチン)神経ホルモンを分泌するペプチド作動性神経細胞が含まれています-神経分泌。 これらの細胞はニューロンと内分泌細胞の両方の特性を持っており、血流から来る信号(ホルモン)と脳内の神経伝達物質と神経ペプチドの両方に反応します。 神経ホルモンはニューロンの細胞質のリボソームで合成され、軸索に沿って末端に輸送されます。
ゴナドトロピン放出ホルモン(リベリン)は、FSHとLHが合成される下垂体のゴナドトロピン機能を調節する神経ホルモンです。 放出ホルモンLH(ルリベリン)が単離され、合成され、詳細に説明されています。 今日まで、卵胞刺激ホルモン放出ホルモン、またはフォリベリンを単離および合成することは不可能でした。
ゴナドリベリンの分泌には脈動する特徴があります。数分間続くホルモンの分泌増加のピークは、比較的低い分泌活性の1〜3時間間隔で置き換えられます。 GnRH分泌の頻度と振幅は、エストロゲンのレベルによって調節されます。
下垂体前葉によるプロラクチンの分泌を制御する神経ホルモンは、プロラクチン阻害ホルモン(因子)またはドーパミンと呼ばれます。
生殖器系の重要なリンクは下垂体前腺です-性腺刺激ホルモン、卵胞刺激ホルモン(FSH、フォリトロピン)、黄体形成ホルモン(LH、ルトロピン)およびプロラクチン(Prl)が機能を調節する腺下垂体卵巣と乳腺は分泌されます。 3つのホルモンはすべてタンパク質物質(ポリペプチド)です。 性腺刺激ホルモンの標的腺は卵巣です。
下垂体の前葉では、甲状腺刺激(TSH)および副腎皮質刺激ホルモン(ACTH)ホルモン、ならびに成長ホルモンも合成されます。
FSHは、卵胞の成長と成熟を刺激し、卵胞顆粒膜細胞の表面でのFSHおよびLH受容体の形成を促進し、成熟卵胞のアロマターゼの含有量を増加させ、芳香化プロセスを刺激することにより、アンドロゲンのエストロゲンへの変換を促進します。卵胞の成長において抑制的かつ刺激的な役割を果たすインヒビン、アクチビン、およびインスリン様成長因子-1の産生を刺激します。
LHは刺激します:
卵胞膜細胞におけるアンドロゲンの形成;
FSHによる排卵;
黄体形成中の顆粒膜細胞のリモデリング;
黄体におけるプロゲステロンの合成。
プロラクチンは、乳腺の成長と授乳を刺激し、黄体のLH受容体の形成を活性化することにより、黄体によるプロゲステロンの分泌を制御します。 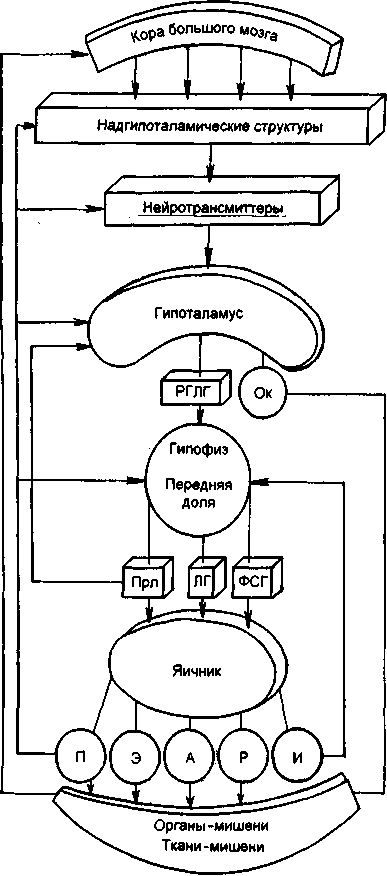
米。 2.14。生殖器系の機能(図)。
RHLH-ホルモンの放出; OK-オキシトシン; Prl-プロラクチン; FSH-卵胞刺激ホルモン; P-プロゲステロン; E-エストロゲン; A-アンドロゲン; Rはリラキシンです。 I-インヒビン; LHは黄体形成ホルモンです。 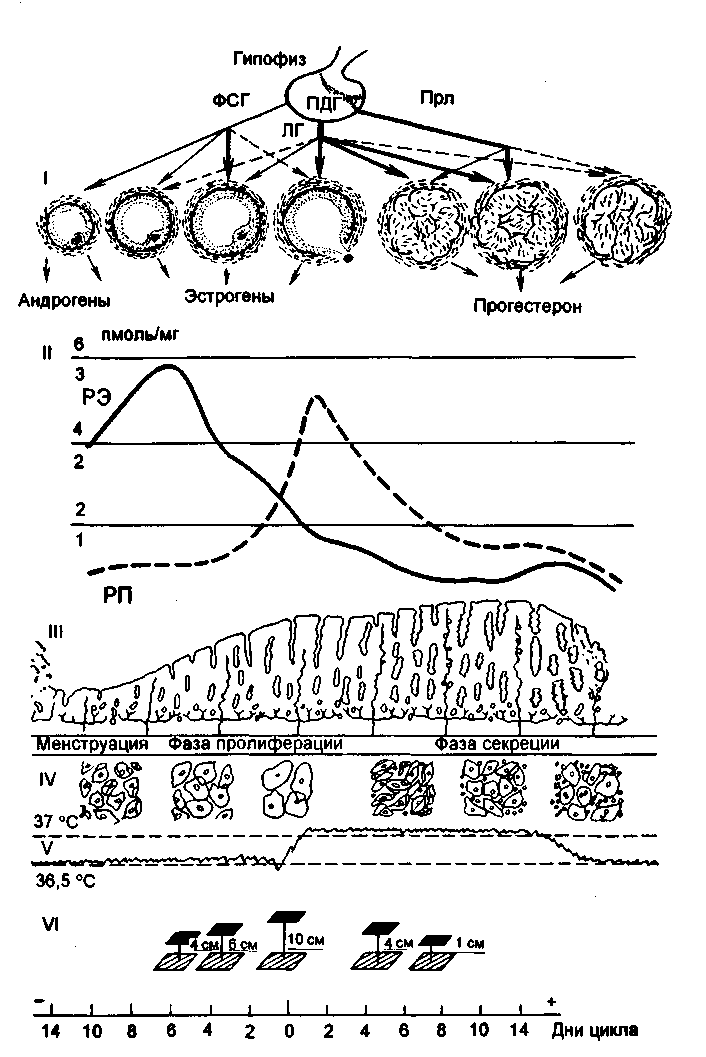
米。 2.15。月経周期中の生殖器系の器官の周期的変化。 I-卵巣機能の性腺刺激ホルモン調節:PDH-下垂体前葉、他の呼称は図と同じです。 2.14; II-エストラジオールの受容体の子宮内膜の含有量-ER(1,2,3;実線)およびプロゲステロン-RP(2,4,6;破線); III-子宮内膜の周期的変化; IV-膣上皮の細胞診; V-基礎体温; VI-頸管粘液の緊張。
下垂体前葉によるプロラクチンの合成は、ドーパミン、またはプロラクチン阻害因子の強壮性遮断制御下にあります。 プロラクチン合成の阻害は、妊娠中、授乳中に停止します。 プロラクチン合成の主な刺激因子は、視床下部で合成されるチロリベリンです。
視床下部-下垂体系と卵巣の周期的変化は相互に関連しており、フィードバックの種類に応じてモデル化されています。
次のタイプのフィードバックが区別されます。
1)フィードバックの「長いループ」-卵巣ホルモンと視床下部の核の間。 卵巣と下垂体のホルモンの間;
2)「短いループ」-下垂体の前葉と視床下部の間。
3)「超短ループ」-GTRHと視床下部の神経細胞の間。
これらすべての構造の関係は、それらの中にある性ホルモン受容体の存在によって決定されます。
生殖年齢の女性は、卵巣と視床下部-下垂体系の間に負と正の両方のフィードバックがあります。 負のフィードバックの例は、下垂体前葉によるLH分泌の増加です。 低レベル周期の初期卵胞期のエストラジオール。 正のフィードバックループの例は、血中のエストラジオールの排卵最大値に応答したLHの放出です。
生殖器系の状態は、基礎体温、瞳孔症状、核分裂指数などの機能診断テストの評価によって判断できます(図2.15)。
基礎体温は、朝、ベッドから出る前に直腸で測定されます。 排卵月経周期では、基礎体温は周期の黄体期に0.4〜0.6°C上昇し、第2期全体にわたって持続します(図2.16)。 月経の当日またはその前日には、基礎体温が低下します。 妊娠中の基礎体温の上昇は、プロゲステロンの影響下での視床下部の体温調節中心の興奮によって説明されます。
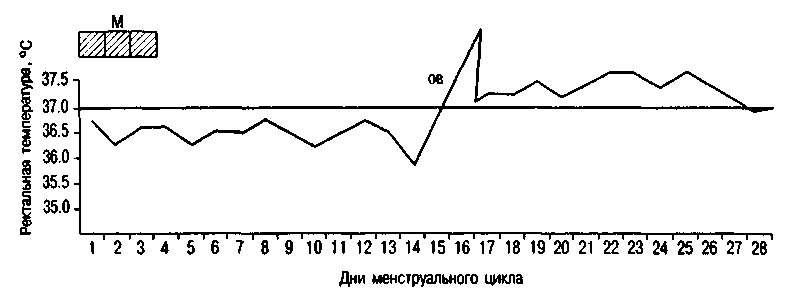
米。 2.16。二相性サイクルにおける直腸温。 M-月経; OV-排卵。
瞳孔の症状は、頸管粘液の変化を反映しています。 エストロゲンの影響下で、透明な硝子体粘液が子宮頸部に蓄積し、それが子宮頸部の外部開口部の拡張を引き起こします。 粘液の最大量は、周期の排卵前の日に観察され、外側の開口部は暗くなり、瞳孔に似ています。 サイクルの第2段階では、プロゲステロンの影響下で、粘液の量が減少するか、完全に消失します。 粘液はゴツゴツした構造になっています。 瞳孔の症状には、+、++、+++の3つの程度があります。
Karyopyknoticインデックス。 卵巣ホルモンの影響下で、周期的な変化は膣粘膜、特にその上部3分の1でも発生します。 膣からの塗抹標本では、扁平上皮の次のタイプの細胞が見られます:a)角質化、b)中間、c)基底、または萎縮。 最初のタイプの細胞は、卵巣によるエストロゲンの分泌が増加するにつれて優勢になり始めます。 細胞要素の定量的比率の決定に基づいて、エストロゲンホルモンによる体の飽和度またはそれらの不足を判断することができます。 角質化細胞の最大数は、排卵前の日に検出されます-80-88%、増殖の初期段階-20-40%、分泌の後期-20-25%。
女性の体内の神経内分泌障害の正しい臨床評価、したがって、それらの病因療法の原理と方法の決定のために、まず第一に、生殖器系の5つのリンクの調節を知る必要があります。その機能は生物種の繁殖です(図1)。
生殖器系機能の調節は、視床下部-下垂体リンクによって決定され、視床下部-下垂体リンクは、神経伝達物質および神経伝達物質を介して大脳皮質によって制御されます(Lakoski J.M.、1989)。
視床下部は、体の一種の生物学的時計、つまり、体の外部および内部環境からの情報を実装し、それによって内部ホメオスタシスを提供する神経調節プロセスの自己調節および自動化のシステムです。生理学的プロセスの通常の過程。 視床下部-下垂体-卵巣複合体の活動を調整する重要なリンクであるのは視床下部であり、その機能はフィードバックメカニズムによってCNS神経ペプチドと卵巣ステロイドの両方によって調節されています(Wildt L.、1989; Sopelak VM、1997)。 。
生殖器系の末梢リンクの現代文学におけるかなり良い照明と、絶えず増加する精神的感情的ストレスの役割の増加を考えると、図1.生殖システムの機能構造ホルモン障害の発症メカニズムについては、参加のいくつかの側面についてより詳細に検討することが適切であると考えました。 視床下部構造 生殖器系の調節において。
ご存知のように、脳は2種類の細胞で構成されています。ニューロンはすべての脳細胞の10%を構成し、グリア細胞はアストロサイトとオリゴデンドライトで、それぞれ残りの90%を構成します。
ニューロンとグリアの発達は、神経上皮前駆細胞(幹細胞)から起こり、その発達の結果として、さまざまな種類のニューロンが発生するニューロン前駆細胞と、後にグリア前駆細胞の2つの細胞株が合成されます。アストロサイトとオリゴデンドロサイトが発達します(Lakoski JM、1989; Sopelak VM、1997)。
ニューロン-これらは、明確なサイズ、形状、細胞小器官を備えた高度に分化した細胞です。 赤血球を除いて、他のすべての細胞と同様に、ニューロンは細胞体を持っており、その中心には核があり、異なる体積の細胞質に囲まれています。
知覚プロセス(樹状突起および唯一の主要な伝達プロセス)は、ニューロンの表面から分岐する軸索であり、特定のシナプス標的細胞まで伸びており、長さが大幅に異なる可能性があります(Sopelak V.M.、1997)。
ニューロンの生命活動の重要なプロセスは、細胞体の細胞質(細胞体とも呼ばれます)に集中し、ニューロン合成の産物が軸索と樹状突起に輸送されます。 細胞体の一部と遠位プロセスの間の両側輸送は、神経機能の完全性を保証し、一定のエネルギー依存の調和のとれたプロセスです。
グリア細胞(英語の単語から のり-接着剤)は当初、脳細胞をサポートすると考えられていましたが、最近の研究では、ニューロンの生命活動の調節におけるそれらの重要な機能的役割が特定されています。 このクラスの非ニューロン細胞要素は、ニューロンの数の9倍であり、実際にはそれらの間の相互作用を保証します。
最も多くのグリア細胞は名前が付けられています アストロサイト、マルチシュートのアウトラインに感謝します。 これらの細胞は、グリア線維性酸性タンパク質の独特な発現を特徴とし、血管、ニューロン、およびそれらの接続の外表面の間に位置しています(図2)。 アストロサイトのプロセスは、ニューロンから毛細血管に向けられ、そこで血管周囲の基盤を形成します。
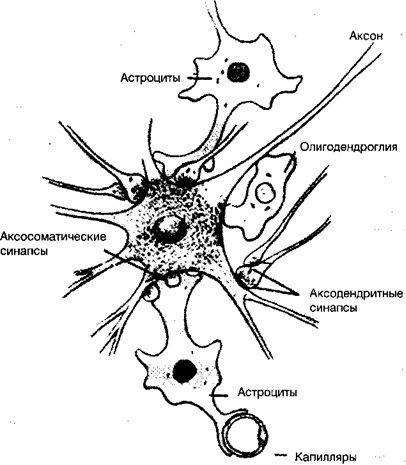 アストロサイトの毛細血管基部は、人間の脳の毛細血管の約85%を覆い、血液脳関門を形成します。
アストロサイトの毛細血管基部は、人間の脳の毛細血管の約85%を覆い、血液脳関門を形成します。
グリア細胞の別のクラスは オリゴデンドロサイト軸索のミエリン鞘を形成する(短い突起と厚い突起の数が少ない細胞)。これにより、ニューロンは神経系内の長距離にわたって弱体化することなく、その効果を迅速に発揮することができます。 オリゴデンドロサイトにはP450ステロイド酵素も含まれており、コレステロールからプレグナノロンを生成します。
脳組織におけるステロイド産生酵素の測定は、生殖機能の調節における中枢神経系の関与のメカニズムの開示に貢献する発見の1つであり、変化の影響下での中枢神経系の変化を説明することも重要です。ホルモンの恒常性において。
アストロサイトにおける神経活性ステロイドの分泌は、オリゴデンドロサイトおよびニューロンよりも高く、これに関連して、これらの細胞の特徴をより詳細に調べる必要があります。
アストロサイトの特性は異なり、まだ十分に研究されていませんが、アストロサイトがニューロンのパラクリン細胞であるという証拠はすでにあります。
アストロサイトでは、インスリン様成長因子(IGF)の存在が明らかになり、その含有量は思春期までに増加し、エストロゲンによる治療中にも増加します。
アストロサイトの一種であるピチサイトは、神経下垂体後葉の主要な非神経細胞要素であり、神経分泌神経終末からのオキシトシンとバソプレッシンの放出を制御する上で重要な役割を果たします。
アストロサイトに黄体形成ホルモン(LH)とヒト絨毛性ゴナドトロピン(CG)受容体が存在することは、LHとCGがグリア細胞の機能に影響を及ぼし、したがって脳の発達と機能に影響を与える可能性があることを示唆しています。
アストロサイトは、インターロイキン(IL-1、IL-2、IL-6)、腫瘍壊死因子a、トランスフォーミング成長因子-ワスプ、インターフェロン、プロスタグランジンEなどのさまざまな免疫調節分子を産生することができ、プロラクチンは有糸分裂誘発とサイトカインを誘導しますアストロサイトでの発現;
アストロサイトは、ニューロンと同様に、脳に広く存在するコルチコトロピン放出因子結合タンパク質(CRF-SP)を産生することができます。 デキサメタゾン、ヒドロコルチゾン、および程度は低いがデヒドロエピアンドロステロンなどのステロイドは、星状細胞からのCRF-SPの放出を阻害します。
視床下部起源の星状細胞は、ニューロンのゴナドトロピン放出ホルモン(Gn-RH)の遺伝子発現を刺激するトランスフォーミング成長因子aおよび(3を分泌しますが、視床下部星状細胞は、デヒドロエピアンドロステロンの合成において皮質の星状細胞よりも約4倍活性があります( DHEA)。
アストロサイトはまた、興奮性効果を提供するグルタミン酸の神経伝達物質レベル、および抗不安(鎮静)効果を達成する上で重要な役割を果たすγ-アミノ酪酸(GABA)の調節に関与する可能性があります。
現在、伝達物質には、アミノ酸、モノアミン、神経ペプチドの3つの主要な化学形態があります。
アミノ酸エキサイティングで気のめいるような送信機として機能します。 伝達物質の興奮性化合物では、アセチルコリン、グルタミン酸、アスパラギン酸が重要です。 抑制性化合物は、GABAやグリシンなどのアミノ酸によって調節されています。
モノアミン、翻訳者として、彼らはカテコラミン作動性(アドレナリン、ノルエピネフリン、ドーパミン)とセロトニン作動性伝達物質で構成されています。 したがって、チロシンは血流からカテコールアミンニューロンに到達し、チロシンヒドロキシラーゼがドーパの合成を触媒する基質です。 ドーパからドーパミンへの変換は、アミノ酸デカルトキシラーゼ(AKD)を介して行われます。 ノルアドレナリン作動性ニューロンのドーパミン-(3オキシダーゼ(DVO)は、ドーパミンをノルエピネフリン(NA)に変換します。
DAとHAはシナプス間隙に放出され、そこでシナプス後受容体にすばやく結合します。 血漿中では、過剰な伝達物質は、カテコール-O-メチルトランスフェラーゼ(COMT)による代謝不活性化、またはシナプス前受容体による再取り込みのいずれかを受け、モノアミノオキシダーゼ(MAO)による代謝分解を受けて、デヒドロキシフェニルエチルグリコール(DOPEthylglycol)を形成します。
ペプチドトランスミッター。視床下部のペプチド含有ニューロンは、もともと神経分泌ニューロンとして説明されていましたが、後に、事実上すべての視床下部神経ペプチドが脳の多くの領域に投射されることが知られるようになりました。 それらは、食物摂取、摂食および性行動の調節において神経伝達物質機能を提供します(表1)。
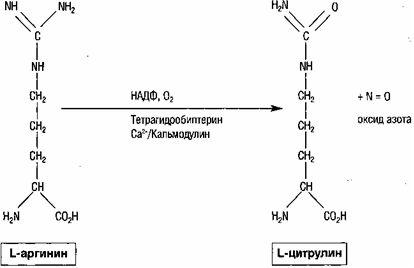
これとは別に、中枢神経系と末梢神経系における一酸化窒素の役割について詳しく説明する必要があります。この発見により、シナプス伝達に関する以前から存在していた見方が根本的に変わりました。 一酸化窒素が神経伝達物質として機能するという実質的な証拠がありますが、それは異常な伝達物質であることに注意する必要があります。 シナプス小胞に貯蔵できない不安定なガスです。 一酸化窒素は、オキシダーゼシンテターゼを使用してL-アルギニンから合成され、他の神経伝達物質のようなエキソサイトーシスではなく、単純な拡散によって神経終末から得られます(図3)。 さらに、一酸化窒素は、他のすべての可逆的神経伝達物質のように受容体と可逆反応を起こしませんが、イソニル酸シクラーゼや他の分子などの酵素を含むいくつかの潜在的な標的と共有結合化合物を形成します。
:可逆性神経伝達物質の作用は、シナプス前放出または酵素分解によって制限されますが、一酸化窒素の作用は、標的からの拡散またはスーパーオキシドアニオンとの共有結合の形成によって媒介されます。
脳内のアルギニンからの一酸化窒素の形成は、補酵素としてNADPHを用いた酸素の存在下でオキシダーゼシンテターゼによって触媒されます補因子としてのテトラヒドロバイオプロテイン。
表1中枢神経系のペプチド伝達物質
(1999年のYen S.S.C.による変更と追加)
|
下垂体ホルモン |
胃腸私ペプチド |
|
甲状腺刺激ホルモン放出ホルモン |
血管作動性腸管ポリペプチド |
|
ゴナドトロピン放出ホルモン |
コリキストキニン |
|
コルチコトロピン放出ホルモンとウロコルチン |
ガストリン |
|
成長ホルモン放出ホルモン |
サブスタンスP |
|
ソマトスタチン |
ニューロテンシン |
|
中枢性尿崩症ホルモン |
メチオニンエンケファリン |
|
1バソプレシン |
ロイシンエンケファリン |
|
オキシトシン |
ダイノルフィン |
|
ニューロフィジン1とN |
ネオエンドルフィン |
|
他の |
インスリン |
|
ニューロペプチドY |
グルカゴン |
|
アンジオテンシンI |
ボンベシン |
|
ブラジキニン |
セクレチン |
|
カルノシン |
ソマトスタチン |
|
カルシトニン遺伝子依存性ペプチド |
モチリン |
|
インターロイキン-1 |
下垂体ペプチド |
|
インヒビン |
副腎皮質刺激ホルモン |
|
パンクレリパーゼ |
p-エンドルフィン |
|
アミリン |
a-メラノサイト刺激ホルモン |
|
副甲状腺ホルモン依存性ペプチド |
プロラクチン |
|
アクチビン |
黄体形成ホルモン |
|
成長ホルモン 甲状腺刺激ホルモン |
生殖器系の中枢調節における一酸化窒素の役割に関して、NOはGn-RHの放出を調節する神経伝達物質であることに注意する必要があります。
神経ステロイド。エストロゲンの局所合成の視床下部での発見(Naftollin et al、1975)は、脳がステロイド産生の機能によって特徴付けられることを示唆しました。 1981年に、プレグナノロンと硫酸プレグナノロン、およびデヒドロエピアンドロステロン(DHEA)とデヒドロエピアンドロステロン硫酸塩(DHEA-S)の存在が、成体の雄ラットの脳で発見されました。 これは、神経ステロイドと呼ばれる中枢神経系におけるステロイド生合成のメカニズムの発見につながりました。
V 人間の脳神経伝達物質のような神経ステロイドは、60歳以上の男性と女性に見られます。 DHEA、プレグナノロン、プロゲステロンは脳のすべての部分に存在しますが、脳内のそれらの濃度は血漿中の数倍です。
脳内では、DHEA-硫酸トランスフェラーゼとスルファターゼの存在も明らかになったため、DHEA-Sの合成は脳内で直接起こっていると考えられます。
組織特異的核内受容体であるステロイド産生因子-1(SF-1)は、いくつかのステロイド産生酵素の遺伝子を調節し、大脳辺縁系の構成要素を含むヒトの脳で広く表されています。
神経ステロイドは、体のすべてのライフプロセスで非常に重要な役割を果たし、GABA受容体、グルタミン酸受容体の活性を調節し、認知機能に影響を与え、神経組織に栄養効果をもたらし(髄鞘形成を促進)、視床下部で放出ホルモンの産生を調節します(Yen SSC、1999)。
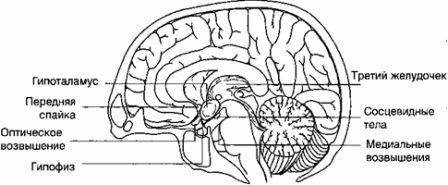
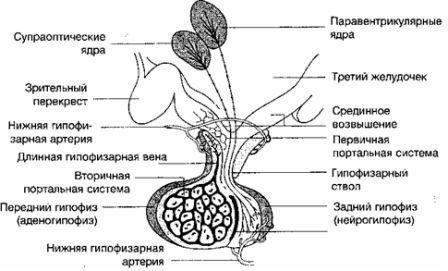
視床下部 -これは、視交叉と正中隆起の間の第三脳室の下にある間脳の一部であり、下垂体幹を介して下垂体の後葉に接続し、対になった乳様体にも接続します(図4 )。視床下部は、さまざまな循環および神経接続を介して中枢神経系および下垂体と相互接続されています。 それは核にグループ化された神経細胞で構成されています。 視床下部の室傍核と視索上核にグループ化された細胞は、バソプレッシン、オキシトシン、ニューロフィジンが放出される下垂体の後葉まで続きます。 この場合、視索上核と室傍核は下垂体の後葉と直接神経接続しています。 視索上核は主にバソプレッシンを分泌し、室傍核はオキシトシンを分泌し、オキシトシンは神経終末に沿って後葉に輸送されます(Sopelak V.M.、1997)。
図H.脳内の一酸化窒素の形成(1999年円S.S.C.)
他の核は放出および抑制因子(Gn-RH、TRH、ソマトスタチン、コルチコトロピン放出ホルモン(CRH))を生成し、これらは循環ポータルシステムを介して下垂体の前葉に輸送され、脳下垂体。
下垂体前葉との機能的接続は、視床下部-下垂体血管系によって表されます(Wildt L.、1989)。 視床下部ホルモンは、正中隆起と視床下部門脈の血流を通って前葉に入ります。 視床下部はまた、視床下部内神経接続、中脳および大脳辺縁系との求心性線維接続、中脳および大脳辺縁系との遠心性線維接続、ならびに下垂体の後葉とを持っています。 視床下部因子は神経線維に沿って正中隆起に輸送され、そこで下垂体毛細血管の壁に浸透します(図5)。 これらの要因は下垂体の内分泌細胞に影響を及ぼし、特定のホルモン反応をもたらします(Yen S.S.C.、1999)。
生殖器系の調節について言えば、視床下部の放出ホルモンの影響下で、性腺刺激ホルモンの合成が下垂体で行われることを強調する必要があります。 化学的性質によりデカペプチドである下垂体放出ホルモン(リベリン)の合成場所は、まさに中基底視床下部の弓状核です。 放出ホルモンの産生は、サーコラルと呼ばれる特定の脈動リズムで起こります。
ゴナドトロピンの正常な分泌を確実にするために、生理学的量のGn-RGの放出の安定した頻度を維持することで十分です。 Gn-RHの放出頻度の変化は、下垂体から分泌されるLHとFSHの量だけでなく、それらの比率も変化させますが、Gn-RHの濃度が10倍に増加しても、FSHはわずかに増加するだけです。分泌物であり、LH分泌物を変化させることはありません(Halvorson LM et al。、1999)。
したがって、リズムの増加は、FSHの放出の有意な増加とLHの放出の減少につながります。 黄体期では、内因性アヘン剤を介したプロゲステロンはパルス発生器の周波数を低下させ、この作用はプロゲステロンの濃度ではなく、その効果の持続時間によって決定されます。 視床下部および性腺刺激ホルモン(Gn-RH受容体の密度の増加)に作用するエストラジオールは、LH / FSH波の振幅を増加させます。
|
|
ヒトにおけるGn-RGの放出の頻度は、70〜90分で1回の放出であり、多くのバイオリズム(睡眠段階の変化、糸球体濾過率と胃液分泌の変動、閉経中のほてりの頻度、 NS。)。 情報の周波数変調は、生殖システムの調節の速度と信頼性、および干渉に対するその耐性を保証します。
パルスリズムジェネレーター-生理学的状態の視床下部の弓状核は、短いフィードバックシステムを介して下垂体によるゴナドトロピンの放出に関する情報を受け取ります。これは、特殊な括約筋が血流のポータルシステムの圧力勾配と血液の一部を調節するためです。下垂体は視床下部に逆流し、視床下部に非常に高い局所濃度の下垂体ホルモンを供給します(Yen S.、1999)。
下垂体におけるLHとFSHの合成と分泌は、同じ細胞によって行われます(Halvorson L.M. et al。、1999)。 性腺刺激ホルモンの表面にはGn-RHの受容体があり、その密度は血中のステロイドホルモンのレベルとGn-RHの濃度に依存します。 Gn-RHと受容体の組み合わせにより、カルシウムイオンが細胞に大量に流入し、数分後にLHとFSHの蓄えが血流に放出されます。 さらに、Gn-RHはLHとFSHの合成を刺激し、性腺刺激ホルモンの完全性を維持します(Wildt L.、1989)。
内分泌腺の機能の調節における重要な役割はに属します 下垂体。それは脳の基部にあるトルコ鞍にあり、前葉(腺下垂体後葉)、中葉および後葉(神経下垂体後葉)で構成されています。 人間の中間的なシェアは事実上存在しません。 下垂体は下垂体幹を介して視床下部に接続します(図5を参照)。
下垂体前葉 5つで構成されています 他の種類免疫学的および超微細構造的特徴が異なる細胞。 前葉のこれらの細胞は、6つの既知のホルモンを産生します。
副腎皮質刺激ホルモン(ACTH)、またはコルチコトロピン;
甲状腺刺激ホルモン(TSH)、またはチロトロピン;
性腺刺激ホルモン:卵胞刺激ホルモン(FSH)、またはフォリトロピン、および黄体形成(LH)、またはルトロピン;
成長ホルモン(STH)、または成長ホルモン;
プロラクチン。
最初の4つのホルモンは、いわゆる末梢内分泌腺の機能を調節し、ソマトトロピンとプロラクチンは標的組織に直接作用します(Halvorson L.M. et al。、1999)。
成長ホルモンとプロラクチンは、好酸性系列に属する成長ホルモン産生細胞とラクトトロピン産生細胞(マンモトロピン産生細胞)の2種類の細胞によって産生されます。 ACTHおよびp-リポトロピンやエンドルフィンなどのプロオピオメラトコルチン分子の他の画分は、甲状腺刺激ホルモンによって合成され、LHおよびFSHは、好塩基性系列に属する性腺刺激ホルモンによって合成されます。
ゴナドトロフは下垂体の前葉の細胞組成の10〜15%を構成し、ラクトトロフの近くにあります。 この局在化の特徴は、これらの細胞の2つのタイプの間にパラが存在することを示唆しています。 緊密な関係(Sopelak V.M.、1997)。
すでに述べたように、前葉のこれらの6つのホルモンの分泌は、視床下部から分泌され、視床下部-下垂体門脈血管を通って下垂体に入る視床下部放出および抑制因子によって制御されます。 しかし、熱帯ホルモンの産生は、生殖器系の中央部(P-エンドルフィン)と末梢部(エストラジオール)の両方で合成される他の物質の影響を受ける可能性があります(Halvorson L.M. et al。、1999)。
下垂体後葉下垂体幹(図5を参照)、神経葉、正中隆起(視床下部の基部にある特別な神経組織で、下垂体調節神経分泌物を下垂体前葉に伝達するための主要な領域を形成します)が含まれます。 下垂体の後葉の2つのホルモン(バソプレッシンとオキシトシン)は、対応するニューロフィジンとともに顆粒に蓄積し、軸索に沿って輸送され、軸索の末端部分に集められ、そこで放出を引き起こす対応するインパルスまで保存されます。 神経ペプチドは、エキソサイトーシスによって分泌顆粒から放出されます。 このプロセスには、神経分泌顆粒の膜と軸索の端にある細胞膜の小さな部分の溶解が含まれます。 顆粒の内容物は細胞間空間に入り、そこから血流に入る(Sopelak V.M.、1997)。
生殖の調節と性腺の機能は、主に下垂体前葉によって分泌される性腺刺激ホルモンによって実行されます。 FSH、LHおよびプロラクチン。 FSH-顆粒膜細胞の増殖を引き起こし、卵胞の成長を刺激します。 LH-アンドロゲンの合成を活性化し、FSHと一緒に排卵を促進します。 FSHと薬物の分泌は、フィードバックメカニズムに従ってゴナドトロピン放出ホルモンによって調節され、エストロゲンとアンドロゲンのレベルにも依存します。 ゴナドリベリン(ルリベリン)は、1時間あたり1パルスから1日あたり1〜2パルスの頻度のパルスによって分泌されます。 ゴナドリベリンの分泌の制御は、性腺刺激ホルモンや他のホルモン、カテコールアミン、アヘン剤ホルモンなどの中枢神経系の多数の神経伝達物質によって行われます。ゴナドリベリンは性腺刺激ホルモンの膜にある受容体と相互作用し、受容体の活性化には存在が必要です最初の3つのアミノ酸の。 ゴナドリベリンのアゴニスト(ブセリリン、ナファレリン、リュープロリドなど)は、同じ膜受容体と相互作用することによってその効果を発揮します(Halvorson L.M.、1999)。
プロラクチンは性腺刺激ホルモンの産生を阻害します。 糖質コルチコイドはまた、LH分泌を抑制する効果があります。
化学構造によれば、LHとFSHは2つのポリペプチドサブユニットaとpからなる糖タンパク質です。 これらのホルモンのa-サブユニットは各糖タンパク質に共通であり、アミノ酸の配列は同じですが、P-サブユニットは糖タンパク質間でアミノ酸の位置の配列が異なります。 ホルモンの特異性に関与するのはPサブユニットです。 両方のサブユニットは、個別に、生物学的に不活性です。 ヘテロダイマーの形成は、生物活性の発現の前提条件です(Halvorson L.M.、1999)。
血中を循環するゴナドトロピンの半減期は、ホルモン分子のシアル酸成分に直接関係しています。 脱シアル化はゴナドトロピンの半減期と生物活性を短縮することが証明されています。 FSHは遊離型で血中にあり、その半減期は55〜60分、LHは25〜30分です。 生殖年齢では、LHの1日あたりの放出量は500〜1100 mIUであり、閉経後、LH形成の速度は増加し、その量は1日あたり最大3000〜3500 mIUです(Sopelak V.M.、1997)。
ステロイドのように、ゴナドトロピンは特定の受容体の活性化を通じて標的組織に生物学的効果を発揮します。 ただし、ステロイドホルモンとは異なり、ゴナドトロピン受容体は標的細胞の膜に関連しています。 ペプチド糖タンパク質ホルモンの細胞表面受容体は、細胞膜の構造の一部であるタンパク質です。 ゴナドトロピンに結合した後、膜受容体は可溶性細胞内メッセンジャーの産生を刺激し、それが次に細胞応答を提供します(Halvorson L.M.、Chin W.W.、1999)。
視床下部リベリンに加えて、現代の概念によると、FSH産生の調節因子は インヒビンとアクチビン、これらは、卵巣顆粒膜細胞と黄体細胞、および細胞栄養芽層細胞によって産生されます(Hopko Ireland et al、1994)。
インヒビンショウブの2つのサブユニットで構成されています。 FSHは、フィードバックの原理に従って、インヒビンの合成と放出に影響を与えます。 oサブユニットと(3サブユニットの組み合わせはFSHの抑制につながり、2つの(3サブユニットの組み合わせは形成につながる) アクチビンしたがって、FSH刺激。
FSHの合成と放出も影響を受けます フォリスタチン、卵胞液から分離されました。 フォリスタチンは、インヒビンと同様に、培養下垂体性腺刺激ホルモン細胞におけるFSHの放出を減少させる糖タンパク質です。 さらに、それはアクチビン結合に対して高い親和性を有し、インヒビン結合に対してはそれほど顕著ではない。 フォリスタチンとアクチビンAは、卵胞のオートクリン-パラクリンシステムの構成要素であり、グラーフ小胞の内膜の細胞のさまざまな機能の調節に関与していることが確立されています(Grome N.、O "Brien ML、1996 )。
ゴナドトロピン分泌には、強壮性、周期的および一時的、または脈動性の3つのタイプがあります(Halvorson L.M.、Chin W.W.、1999)。
ゴナドトロピンの強壮性または基礎的な分泌は、負のフィードバックによって調節され、周期的-エストロゲンの関与を伴う正のフィードバックメカニズムによって調節されます。
脈動する分泌物は、視床下部の活動とゴナドリベリンの放出によるものです。
サイクルの前半での卵胞の発達は、FSHとLHの強壮性分泌によるものです。 エストラジオールの分泌の増加は、FSHの形成の阻害につながります。 卵胞の発達は、顆粒膜ゾーンの細胞内のFSH受容体の数に依存し、これらの受容体の合成は、エストロゲンによって刺激されます。
こちらです、 FSH特定の卵胞でエストロゲンが合成され、FSHの受容体の数が増えることで、その蓄積(受容体に結合することで)、卵胞のさらなる成熟、エストラジオール分泌の増加に寄与します。 この時点で他の卵胞は閉鎖状態になります。 血中のエストラジオールの濃度は排卵前の期間に最大に達し、それは大量のゴナドリベリンの放出とそれに続くLHとFSHの放出のピークにつながります。 LHとFSHの排卵前の増加は、グラーフ小胞の破裂と排卵を刺激します(Hurk Van Den R.、1994)。
LH卵巣におけるステロイド合成の主な調節因子です。 LH受容体は黄体細胞に局在し、LHの効果は、アデニル酸シクラーゼの刺激と、直接または中間体(プロテインキナーゼなど)を介してプロゲステロンに関与する酵素を活性化するcAMPレベルの細胞内増加によって媒介されます。生合成。 卵巣のLHの影響下で、ホルモンの合成に必要なコレステロールの量が増加します。 同時に、コレステロール分子の側鎖を切断するシトクロムP450ファミリーの酵素の活性が増加します。 より長い曝露で、LHはプロゲステロンと他のステロイドの合成に関与する他の酵素(ZV-ヒドロキシステロイドデヒドロゲナーゼ、17a-ヒドロキシラーゼ)の発現と合成を刺激します。 したがって、黄体では、LHの影響下で、コレステロールがプレグナノロンに変換される部位でステロイド産生のプロセスが促進されます(Yen S.、1999)。
ゴナドトロピンの分泌の調節は、「短い」および「超短い」フィードバックループによって提供されます。 したがって、LHとFSHのレベルの増加は、それらの合成と放出の阻害につながり、視床下部におけるゴナドリベリンの濃度の増加は、その合成と下垂体の門脈系への放出を阻害します(Sopelak V.M.、1997)。
GnRHの放出は、カテコールアミンの影響も受けます。 ドーパミン、アドレナリン、ノルエピネフリン。エピネフリンとノルエピネフリンはゴナドリベリンの放出を刺激しますが、ドーパミンは以前にステロイドホルモンを注射された動物でのみ同じ効果があります。 コレシストキニン、ガストリン、ニューロテンシン、オピオイド、およびソマトスタチンは、ゴナドリベリンの放出を阻害します(Yen S.、1999)。
副腎皮質刺激ホルモン副腎皮質に刺激効果があります。 タンパク質合成の増加(cAMP依存性の活性化)により、副腎皮質の過形成が起こります。 ACTHは、コレステロールの合成とコレステロールからのプレグナノロンの形成速度を高めます。 その効果は、グルココルチコイドの形成の増加につながるバンドルゾーンでより多く発現しますが、糸球体および網状ゾーンではより少ない程度で発現します。したがって、鉱質コルチコイドと性ホルモン。
ACTHの副腎外効果は、脂肪分解を刺激し(脂肪貯蔵から脂肪を動員し、脂肪の酸化を促進する)、インスリンとソマトトロピンの分泌を増加させ、筋肉細胞におけるグリコーゲンの蓄積、インスリン分泌の増加に関連する低血糖を増加させることです。メラノフォアの色素細胞に対する作用による色素沈着。
成長ホルモン成長と身体の発達の調節に関与し、体内のタンパク質の形成、RNA合成、および血液から細胞へのアミノ酸の輸送に刺激効果をもたらします。
主な生物学的役割 プロラクチン-乳腺の成長と授乳の調節。 これは、タンパク質(ラクトアルブミン、乳脂肪、炭水化物)の合成を刺激することによって行われます。 プロラクチンはまた、黄体の形成とそれによるプロゲステロンの生成を調節し、体の水塩代謝に影響を与え、体内の水分とナトリウムを保持し、アルドステロンとバソプレッシンの効果を高め、炭水化物からの脂肪の形成を増加させます。
下垂体後葉ホルモン視床下部で形成されます。 それらの蓄積は下垂体後葉で起こります。 視床下部の視索上核と室傍核の細胞では、オキシトシンと抗利尿ホルモンが合成されます。 合成されたホルモンは、ニューロフィジン担体タンパク質を使用した軸索輸送によって、視床下部-下垂体を介して下垂体の後葉に輸送されます。 ここでは、ホルモンが沈着し、その後血中に放出されます。
抗利尿ホルモン(ADH)、またはバソプレッシンは、体内で2つの主要な機能を持っています。 その抗利尿作用は、遠位ネフロンでの水の再吸収を刺激することです。 この作用は、ホルモンと特定の受容体との相互作用によって行われ、尿細管壁の透過性、再吸収、および尿の濃度の増加につながります。 この場合、尿細管の細胞内のヒアルロニダーゼの活性化により、水の再吸収の増加も起こり、解重合の増加につながります ヒアルロン酸、その結果、循環液の量が増加します。
大量投与(薬理学的)では、ADHは細動脈を収縮させ、その結果、動脈圧が上昇します。 したがって、それはバソプレッシンとも呼ばれます。 血中の生理的濃度により、この作用は必須ではありません。 失血、痛みのショックで発生するADHの放出の増加は、血管収縮を引き起こし、これらの場合、適応値があります。
ADHの産生の増加は、細胞外液と細胞内液の量の減少、減少とともに起こります 血圧、レニン-アンギオテンシンおよび交感神経系の活性化に伴う浸透圧血圧の上昇。
オキシトシン子宮の平滑筋に選択的に作用し、出産時に収縮させます。 このプロセスは、細胞の表面膜にある特別なオキシトシン受容体に結合することによって実行されます。 高濃度のエストロゲンの影響下で、オキシトシンに対する受容体の感受性が急激に増加します。これは、出産前の子宮の収縮活動の増加を説明しています。
授乳過程へのオキシトシンの関与は、乳腺の筋上皮細胞の収縮を増加させることであり、それにより乳汁の排泄が増加します。 次に、オキシトシンの分泌の増加は、母乳育児中の子宮頸部の受容体、および乳房の乳首の機械受容器からのインパルスの影響下で発生します。
生殖システムの次のレベルは 卵巣、ステロイドと卵胞形成は、ゴナドトロピンの周期的分泌に応答して、成長因子(RF)の影響下で発生します。
卵巣は女性の生殖器系の対の器官であると同時に内分泌腺です。 卵巣は2つの層で構成されています:ツニカアルブギネアで覆われた皮質物質と延髄です。 これとは別に、卵巣アンドロゲンの産生に関与する顆粒細胞を含む、間質黄体細胞を欠く卵巣門のセクションが考慮されます。
皮質物質は、結合組織間質に位置するさまざまな成熟度(原始から閉鎖まで)の濾胞によって形成されます。
卵胞形成のプロセスは卵巣で継続的に起こり、卵巣受容体との相互作用によってゴナドトロピンによって調節されます(Sopelak V.M.、1997)。
同時に、各卵巣で数十個の卵胞が検出され、それらは成長と成熟のさまざまな段階にあります。 出生時の卵胞の総数は約200万個です。月経周期が確立するまでに卵胞の数は8〜10倍減少し、3万〜4万個を超えません。卵胞の約10%のみが排卵の前兆であり、黄色い体に変わります。 残りは閉鎖と退行を経験します(Hurk Van Den R. et al。、1994)。
一次卵胞が成熟卵胞に変化する間に、減数分裂の最初の分裂が完了し、その結果、一方向(極)体が放出され、卵母細胞が形成されます。 透明な膜は最大の発達に達し、無秩序に横たわる濾胞細胞の1〜2層で覆われた放射状の冠に変わります。 卵胞に空洞が形成され、排卵前に最大サイズに達します。 間質の血管の成長因子の影響下にある濾胞細胞の層は、濾胞の内側と外側の流れの2つの層に変わります。 卵胞液の量がさらに増えると、卵胞腔のオーバーフローとその破裂、つまり排卵につながります。 排卵後、放射性の冠に囲まれた卵母細胞は、腹腔から卵管の漏斗に入り、次にその内腔に入ります。 ここで減数分裂の第2分裂が完了し、成熟した卵子が形成され、受精の準備が整います(Yen S.、1999)。
卵巣周期には2つの段階があります — 卵胞期および黄体期 排卵と月経によって分離される遠吠え。
V 卵胞段階下垂体から分泌されるFSHの影響下で、さまざまな成長因子とともに、1つまたは複数の原始卵胞の成長と発達、および顆粒膜細胞の分化と増殖が刺激されます。 FSHはまた、17-(3-ヒドロキシステロイドデヒドロゲナーゼとアロマターゼ、これらはcAMPの活性化を通じて顆粒膜細胞でのエストラジオールの形成に必要であり、したがって一次卵胞の成長と発達、卵胞によるエストロゲンの産生を刺激します)の活性を増強します上皮細胞。これにより、FSHの作用に対する顆粒膜細胞の感受性が高まります。FSH受容体は、7つの膜貫通断片を持つ膜受容体のグループに属します。エストロゲンとともに、少量のプロゲステロンが分泌されます。多くの卵胞のうちそれが成長し始めると、1つだけが最終的な成熟に達しますが、それほど頻繁ではありません-2-3。
ゴナドトロピンの排卵前の放出は、排卵過程自体を決定します。 卵胞の体積は、多形核白血球によって分泌されるタンパク質分解酵素およびヒアルロニダーゼの活性の増加に関連する卵胞壁の薄化と並行して急速に増加します。
排卵前の2〜3日以内に観察されたエストロゲンレベルの有意な増加は、卵胞液の放出を伴う多数の成熟卵胞の死によるものです。 負のフィードバックメカニズムによる高濃度のエストロゲンは、下垂体によるFSHの分泌を阻害します。 LHの排卵放出、および程度は低いがFSHは、超高濃度のエストロゲンおよびLHレベルの正のフィードバックメカニズムの存在、および排卵前の24時間のエストラジオールレベルの急激な低下に関連しています。 。
月経周期の神経ホルモン調節を図6に模式的に示します。
排卵
図6.月経周期の神経ホルモン調節
排卵 LHまたは絨毛性ゴナドトロピンの存在下でのみ発生します。 さらに、FSHとLHは卵胞発育の期間中に相乗剤として作用し、この時点で卵胞膜細胞は活発にエストロゲンを分泌します。
卵胞壁のコラーゲン層の破壊のメカニズムは、卵胞期の妥当性に基づくホルモン依存性のプロセスです。 LHの排卵前の放出は、排卵時のプロゲステロンの濃度の増加を刺激します。 プロゲステロンの最初のピークにより、卵胞壁の弾力性が増加し、FSH、LH、およびプロゲステロンが共同でタンパク質分解酵素の活性を刺激します:顆粒細胞によって分泌されるプラスミノーゲン活性化因子はプラスミンの形成を促進し、プラスミンはさまざまなコラゲナーゼ、プロスタグランジンEを生成しますF 2 otは、卵母細胞塊の蓄積の変位に寄与します。 非排卵卵胞の早期黄体形成を起こさないためには、一定量のアクチビンが卵巣で産生されなければなりません(Speroff L. et al。、1994)。
排卵後、血清LHおよびFSHレベルが急激に低下します。 サイクルの第2フェーズの12日目から、血中のFSHのレベルが2〜3日増加し、新しい卵胞の成熟が始まります。一方、第2フェーズ全体でのLHの濃度はサイクルは減少する傾向があります。
排卵した卵胞の空洞は崩壊し、その壁はひだに集まります。 排卵時の血管の破裂により、排卵後の卵胞の空洞に出血が起こります。 結合組織の瘢痕-汚名は、将来の黄色い体の中心に現れます(Speroff L. et al。、1994)。
LHの排卵放出とそれに続く5〜7日間の高レベルのホルモンの維持は、ルテイン細胞の形成を伴う顆粒膜細胞(顆粒膜)の増殖と腺変態のプロセスを活性化します。 来る 黄体期 (黄体の相) 卵巣周期 (Erickson G. F.、2000)。
毛包の顆粒層の上皮細胞は集中的に増殖し、リポクロームを蓄積して黄体細胞に変わります。 膜自体は豊富に血管新生されています。 血管新生の段階は、顆粒膜上皮細胞の急速な増殖とそれらの間の毛細血管の集中的な成長によって特徴付けられます。 血管は、排卵後の卵胞の空洞に側面から侵入します thecae internae半径方向の黄体組織に。 黄体の各細胞には毛細血管が豊富に供給されています。 結合組織と血管は、中央の空洞に到達し、それを血液で満たし、後者を包み込み、黄体細胞の層からそれを制限します。 黄体は、人体で最も高いレベルの血流を持っています。 この独特な血管網の形成は、排卵後3〜4日以内に終了し、黄体機能の全盛期と一致します(Bagavandoss P.、1991)。
血管新生は、既存の基底膜の断片化、内皮細胞の移動、およびマイトジェン刺激に応答したそれらの増殖の3つの段階で構成されます。 血管新生活性は、線維芽細胞成長因子(FGF)、上皮成長因子(EGF)、血小板成長因子(PRT)、インスリン様成長因子-1(IGF-1)、および次のようなサイトカインなどの主要な成長因子によって制御されます。腫瘍壊死因子(TNF)およびインターロイキン(IL-1; IL-6)(Bagavandoss P.、1991)。
この時点から、黄体はかなりの量のプロゲステロンを生成し始めます。 プロゲステロンは一時的に正のフィードバックメカニズムを不活性化し、ゴナドトロピンの分泌はエストラジオールの負の効果によってのみ制御されます。 これにより、黄体の相の途中でゴナドトロピンのレベルが最小値に低下します(Erickson G.F.、2000)。
黄体の細胞によって合成されるプロゲステロンは、新しい卵胞の成長と発達を阻害し、受精卵の導入のための子宮内膜の準備にも関与し、子宮筋層の興奮性を低下させ、エストロゲンの影響を抑制します周期の分泌期の子宮内膜は、脱落膜組織の発達と乳腺の肺胞の成長を刺激します。 血清プロゲステロン濃度のプラトーは、直腸(基礎)温度(37.2〜37.5°C)のプラトーに対応します。これは、発生した排卵を診断する方法の1つであり、黄体期の有用性を評価するための基準です。 基礎体温の上昇は、プロゲステロンの影響下での末梢血流の減少に基づいており、これにより熱損失が減少します。 血中の含有量の増加は、排卵の指標である基礎体温の上昇と一致します(McDonnel D.P.、2000)。
エストロゲンの拮抗薬であるプロゲステロンは、子宮内膜、子宮筋層、および膣上皮におけるそれらの増殖効果を制限し、子宮内膜腺によるグリコーゲンを含む分泌物の分泌の刺激を引き起こし、粘膜下組織の間質を減少させます。 受精卵の着床に必要な子宮内膜の特徴的な変化を引き起こします。 プロゲステロンは子宮の筋肉の緊張を低下させ、それらをリラックスさせます。 さらに、プロゲステロンは乳腺の増殖と発達を引き起こし、妊娠中は排卵過程の抑制に寄与します(O "Malleu B.W.、Strott G.A.、1999)。
卵胞発達のこの段階の期間は異なります:受精が起こらなかった場合、受精した卵が子宮内膜に侵入し、結果として生じる胞胚が絨毛性ゴナドトロピン(CG)を合成し始めた場合、10〜12日後に月経黄体の退行が起こります)、その後黄色の体は妊娠の黄体になります。
黄体の顆粒膜細胞は、出産時に重要な役割を果たすポリペプチドホルモンのリラキシンを分泌し、骨盤靭帯の弛緩と子宮頸部の弛緩を引き起こし、子宮筋層でのグリコーゲンの合成と水分保持を増加させます。その収縮能力を低下させます。 通常の月経周期の間、その分泌はLH放出のピークの直後に上昇し、月経中も検出可能なままです。 妊娠中、リラキシンの循環レベルは、第2および第3トリメスターと比較して、第1トリメスターの終わりに高くなります。
卵子の受精が起こらなかった場合、黄体は逆発達の段階に入り、月経を伴います。 黄体細胞は、核濃縮が観察される一方で、ジストロフィーの変化、サイズの減少を経験します。 崩壊する黄体細胞の間で成長する結合組織がそれらに取って代わり、黄体は徐々に硝子体に変わります-白い体 (白体)(Sopelak V.M.、1997)。
ホルモン調節の観点から、黄体の退行期間は、プロゲステロン、エストラジオール、およびインヒビンAのレベルの顕著な減少によって特徴付けられます。インヒビンAのレベルの減少は、下垂体およびFSHに対するその遮断効果を排除します。分泌。 同時に、エストラジオールとプロゲステロンの濃度の漸進的な減少は、Gn-RH分泌の頻度の急速な増加を促進し、下垂体は負のフィードバックの阻害から解放されます。 インヒビンAおよびエストラジオールレベルの減少、ならびにGn-RH分泌のインパルスの頻度の増加は、LHよりもFSH分泌の優位性を保証します。 FSHレベルの増加に応じて、胞状卵胞のプールが最終的に形成され、そこから将来的に優勢な卵胞が選択されます。 プロスタグランジンF2 a、オキシトシン、サイトカイン、プロラクチンおよびラジカル0 2は黄体融解効果があり、これが黄体欠乏症の発症の基礎となる可能性があります。 炎症過程付属物で。
卵巣(月経)周期の期間は通常21日から35日まで変化します。
月経は黄体の退行を背景に起こります。 それの終わりまでに、エストロゲンとプロゲステロンのレベルは彼らの最小に達します。 このような背景に対して、視床下部と下垂体の緊張性中心の活性化と、卵胞の成長を活性化する主にFSHの分泌の増加があります。 es-tradolのレベルの増加は、子宮内膜の基底層での増殖過程の刺激につながり、子宮内膜の適切な再生を確実にします(図7)。
図7.通常の月経周期の調節のリンク(Sopelak V.、1997)
卵巣ステロイド産生卵胞の空洞を裏打ちする上皮の細胞、内卵胞膜の細胞、そして間質でははるかに少ない細胞を通過します。 濾胞上皮細胞、間質およびテカ組織は、プロゲステロン、テストステロン、デヒドロテストステロン、エストロンおよびエストラジオールを合成します(Erickson G.F.、2000)。
エストロゲンは、エストラジオール、エストロン、エストリオールに代表されます。 生物学的には、エストラジオールが最も活性が高く、その95%が卵胞で形成され、血中のそのレベルは卵胞の成熟の指標です。 エストラジオール(E 2)は、主に顆粒膜細胞から分泌され、少量では黄体から分泌されます。 エストロン(E、)は、エストラジオールの末梢芳香族化によって形成されます。 エストリオール(E 3)の主な供給源は、肝臓でのエストラジオールとエストロンのヒドロキシル化です(O "Malleu B.W.、Strott G.A.、1999)。
血中に分泌されるエストロゲンは、性ステロイド結合グロブリン(SSSH)によって結合され、程度は低いですが、血中アルブミンによって結合されます。 CCCCは、エストラジオール-テストステロン結合グロブリンとも呼ばれます。 名前自体は、アンドロゲンに対するこのタンパク質の親和性の増加を示しています。 女性の血清中の性ホルモンに結合するグロブリンのレベルは、男性の血中の濃度のほぼ2倍です。 エストロゲンとその代謝物は、肝臓でグルクロン酸と硫酸と結合し、胆汁と尿に排泄されます(McDonnel D.P.、2000)。
性器、下垂体、視床下部に対するすでに述べた効果に加えて、エストロゲンは同化作用を持ち、代謝を高めます 骨組織そして、一方では思春期の開始時の成長の停止に関連する骨格の骨の成熟を加速し、他方では性的発達が遅れた少女の若年性骨粗鬆症の発症を加速します。
大量投与では、エストロゲンは浮腫の発症まで体内のナトリウムと水分の保持を促進します。 それらはまた血中コレステロールレベルを下げることによって脂質代謝に影響を及ぼします。
プロゲステロンは、黄体、副腎皮質、精巣から分泌され、コルチコステロイドやアンドロゲンの生合成の前駆体として使用されます。 プロゲストゲンと糖質コルチコイドは類似した化学構造を持っているので、プロゲステロンと糖質コルチコイド受容体は架橋特性を持っています。 血清中、プロゲステロンはトランスコルチンによって結合されます。トランスコルチンは糖質コルチコイドにも結合することが知られています。 いくつかの研究によると、トランスコルチンに結合するプロゲステロンの能力は、コルチコステロイドの能力を超えています。 肝臓では、プロゲステロンはグルクロン酸に結合し、抱合状態で尿中に排泄されます(McDonnel D.P.、2000)。 ただし、エストロゲンとプロゲステロンが標的臓器に及ぼす影響については、「臨床診療における性ステロイドホルモンの使用の原則とその全身的影響」のセクションで詳しく説明しています。
女性のアンドロゲンは、主にアンドロステンジオンの形で卵巣間質細胞から分泌され、副腎では卵巣の3倍の量で形成されます。 末梢組織のアンドロステンジオンはテストステロンに変換されます。 卵巣では、テストステロン、ジヒドロテストステロン、デヒドロエピアンドロステロンも少量生成されます。 女性の体内で分泌されるテストステロンの約1/4は卵巣で生成されます。 その量の残りは副腎によって分泌されるか、アンドロステンジオンからの変換によって周辺の組織で形成されます(McDonnel D.P.、2000)。
標的組織におけるステロイドの生物学的効果は、それらの特定の受容体の存在と関連しています(図8)。 ステロイドは拡散によって細胞膜を通過し、細胞質内の特定の受容体に結合します。 ステロイド受容体は、特定のホルモンに対する高い結合能力を持つ比較的大きなタンパク質です。 ただし、これらの受容体がこのグループの他のステロイドと結合する可能性があります(たとえば、合成アゴニストおよびアンタゴニスト)。 細胞質受容体はすべてに存在するわけではなく、このタイプのホルモンに敏感な組織細胞にのみ存在します。 受容体-ステロイド複合体は、その形成が温度を含むいくつかの要因に依存し、核に移動します。核には、これらの複合体に結合するクロマチン上の特別な領域があります。 受容体-ステロイド複合体が活性化された後、DNA上にあるアクセプター核タンパク質に結合することができます。 後者の相互作用は、多数の特定のRNAおよび対応するタンパク質の合成、対応する器官(乳腺、子宮など)および組織の成長および発達につながります(O "Malleu B.W.、Strott G.A.、1999)。
|
|
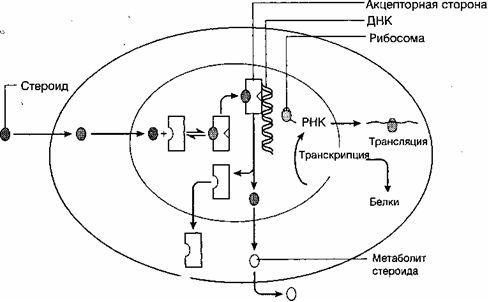
さまざまなステロイドホルモンの受容体分子の数は、細胞あたり5,000から20,000までさまざまです。 エストロゲン受容体は、同じ親和性で多くの天然および合成のエストロゲンステロイドに結合します。 エストロゲンとプロゲステロンの受容体は2つのサブユニットであり、それぞれがホルモン分子に結合すると考えられています。これについては、臨床診療のセクションで詳しく説明します。
aおよびPサブユニットのそれぞれはクロマチンと相互作用し、特定の遺伝子およびRNAポリメラーゼのさらなる活性化を提供します。ホルモンの生物学的効果は、血清中の量的変動だけでなく、受容体リンクの状態にも関連しており、受容体の数は大きな変動の影響を受けます。
|
図8.標的組織に対するステロイドホルモンの作用機序(Cowan B.D.、1997) |
実験的研究は、新生児ラットにおいて、標的組織が少量のエストロゲン受容体を含むことを示しました。 生後10日目に受容体の数が増加し、この期間の後、外因性エストロゲンの導入はそれらの増加を引き起こします。 エストロゲンは、エストロゲンだけでなくプロゲステロンの受容体の形成を刺激します。 受容体の数は、血中を循環するホルモンのレベルに依存するだけでなく、遺伝的制御下にあります。 したがって、アンドロゲン受容体の完全な欠如は、精巣女性化症候群で観察されます(McDonnel D.P.、1999)。
|
|
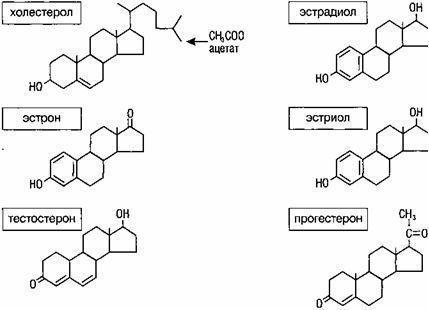
主な性ステロイドホルモンの化学構造の分析は、それらがすべてプロゲステロンの誘導体であり、エストロゲンはそれらの構造に存在するヒドロキシラジカルの数だけが互いに異なることを示しています(図9)。
すべてのステロイドホルモンの物質は、低密度リポタンパク質コレステロール(LDL)です。 ゴナドトロピン(FSHおよびLH)、ならびに酵素系(アロマターゼ)は、ステロイド産生に関与しています。 まず、コレステロール側鎖の切断の結果としてプレグナノロンが形成されます。 将来的には、プレグナノロンの代謝変換の2つの可能な経路があり、テストステロンの形成で終わります。テストステロンは、得られた化合物の不飽和二重結合の位置によって代謝のm-およびl5-経路の名前を受け取りました。
性ステロイドの主な形成は、L5経路に沿って発生します。
|
図9.ステロイドホルモンの化学構造(Sopelak V.、1997) |
その過程で、17-ヒドロキシプレグナノロン、デヒドロエピアンドロステロン(DHEA)、アンドロステンジオンが順次形成されます。
プロゲステロン、17a-ヒドロキシプロゲステロン、アンドロステンジオンは、L4経路に沿って形成されます。 両方の経路A4,5-イソメラーゼを閉じます。 さらに、テストステロンまたはアンドロステンジオンの芳香族化は、それぞれエストラジオールまたはエストロンの形成とともに起こります(図10)。
ノート: HSD-Zp-ヒドロキシステロイドデヒドロゲナーゼ、DOK-デオキシコルチコステロン
図10.ステロイドの生合成(Cowan B.D.、1997)
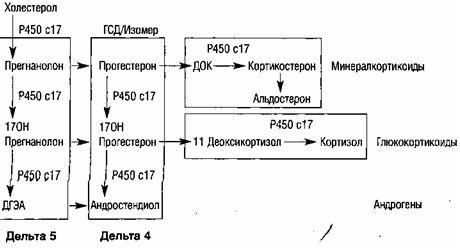
コレステロールを前駆体および生物学的に活性なステロイドに変換するステロイド産生酵素のほとんどは、シトクロムP450グループに含まれています。 シトクロムP450は、多くの酸化酵素の総称です(Bryan D.、1997)。 シトクロムには約200種類あり、そのうち5種類がステロイド産生の過程に関与しています(表2)。
プロセスに関与するP450酵素ステロイド産生
|
NS450 - 新しい名前 |
アロマターゼ |
したがって、生殖システムは単一の統合システムであり、そのすべてのリンクは直接とフィードバックの両方のメカニズムによって相互接続されています。
ナレッジベースであなたの良い仕事を送るのは簡単です。 以下のフォームを使用してください
知識ベースを研究や仕事に利用している学生、大学院生、若手研究者はあなたにとても感謝しています。
ポストする http://www.allbest.ru/
生殖器系の解剖学と生理学
1. 人間の生殖システムは、生殖、性決定、性機能の機能を果たします。 男性で構成されています kihまたは女性の生殖器
1.1男性の生殖器系の解剖学と生理学
男性用生殖器系は、外性器と内性器によって形成され、一次および二次の男性の性的特徴によって特徴付けられます。
外部の男性生殖器陰茎と陰嚢によって形成されます。
陰茎(ラテン語-陰茎;男根から)2つの海綿体と1つの海綿体で形成されています。 頭、体、根があります。 陰茎の皮膚は薄く、脂肪層がなく、簡単にずれます。 尿道(尿道)は海綿体を通り抜けます。 海綿体の組織は、線維弾性組織と平滑筋線維によって形成されたクロスバーで構成されています。 ラングは血で満たされた洞窟を形成します。 陰茎の成長には個々の特徴があり、18〜24年で終わります。 穏やかな状態では、その長さは7〜12 cm、厚さは2〜3 cmです。性的興奮の間、血管の特別な配置により、動脈を通って流れるよりも多くの血液が陰茎に運ばれます。静脈。 これは海綿体の血液の保持につながり、陰茎は密になります(勃起)。 陰茎の頭はと呼ばれる皮膚のひだで覆われています 包皮..。 頭の下側では、包皮は包皮によって頭の皮膚に接続されています。 頭と包皮の間にスペースが残ります-包皮の空洞は、包皮が引き戻されたときに頭が通過できるようにする開口部で正面に開きます。 頭の突き出た尾根の周りには、恥垢と呼ばれる秘密を分泌する腺があります。 恥垢は酸化して、特徴的な臭いのある白い塊を形成します。 陰茎の頭には、尿道の外部開口部が開いています。これは、最大18 cmの長さで、狭く、S字型に湾曲しています。
陰茎機能:男性の性別指標; 尿と精液の排泄; 出産; セクシー。
陰嚢は筋皮神経の嚢状の形成であり、付属肢のある精巣が含まれています。 陰嚢の皮膚は薄く、伸びやすく、体の他の部分よりも色が濃いです。 陰嚢機能:男性の性別指標; 睾丸のためのレセプタクル; 精子の成長と発達および性ホルモンの合成のための条件を確実にするための精巣温度の正常化; 美的; セクシー。 男性の内臓精巣とその付属肢(精管、精嚢、クーパー腺、前立腺)によって形成されます。
睾丸(ラテン語-精巣、ギリシャ語-蘭から)、または男性の性腺は、陰嚢にある対の器官を指します。 睾丸は長さ3.5〜4cm、幅2.5〜3cmの楕円形で、重さは15〜25gです。左の睾丸は低く、通常は右の睾丸よりも低くなっています。 精巣実質は、精巣上体管に通じる精細管で構成されています。 精索は睾丸の後縁に近づきます。 胚の睾丸は腹腔内で発達し、出生時までに鼠径管を通って陰嚢に下降します。 睾丸は思春期に活発に成長し、視床下部-下垂体系からのホルモンの影響下で機能します。 精細管は5mの長さで、精子の生産場所です。 それらは輸精管に分類されます。
精巣機能:男性の性別指標; 男性ホルモンの合成と分泌-アンドロゲン(主にテストステロン); 少量の女性ホルモン(エストロゲンとプロゲステロン)の合成と分泌; 精子の形成と成長; 精子を構成する体液の合成と分泌; セクシー。
精子または男性の生殖細胞は、下垂体ホルモンの影響下で精細管の細胞変換の複雑なプロセスの結果として、72〜74日以内に形成および発達します。 精子細胞は、若い男性の思春期に精巣によって生成され始めます。 彼らの最大の教育は20-30歳になります。 40年後、精子の生産が減少します。 しかし、精子形成は時々老年まで続くことができます。 人間の精子はオタマジャクシに似ています。 それは頭、首、そして長く細い尾を持ち、それは終わりの糸またはそれらの束で終わります。 精子の全長は50〜60ミクロンです。 精子の頭は原形質の薄い層に囲まれた細胞核であり、首と尾は細胞質で構成されています。 精子の核には、遺伝情報がコードされているDNAに染色体の半数体セット(23染色体)が含まれています。 精子は2〜3mm /分の速度で尾を曲げることによって動きます。 精子は、精嚢や前立腺の分泌物と接触すると運動性になります。 顕微鏡で一滴の精液を観察すると、精子の動きが不規則であることに気付くでしょう。 膣、子宮頸管、子宮腔、および卵管の生理学的環境では、精子の動きが命令され、これらの器官の分泌物の流れに逆らって方向付けられます。 3〜4分後、精子は膣から子宮頸管に浸透し、60〜90分後に子宮腔に到達し、2〜3時間後に顔面神経管に入ります。 子宮と卵管のアルカリ性環境では、精子は2〜3日間移動して受精する能力を保持します。 精子の運動性と出産性は、0°Cから40°Cの温度で維持されます。 ある温度で凍結する場合 液体窒素(-192°C)精子は、長期間移動して受精する能力を保持します。 これは、体外受精の目的で精子バンクに精子を保存するために使用されます。 長い間睾丸にあった精子は死に、吸収されて新しいものと交換されます。
精子機能:女性の生殖細胞と融合して接合子を形成し、そこから胎児が発達して成長します。 子孫への遺伝形質の伝達。
前立腺(ラテン語-前立腺から)形状は直径4cmの栗の実に似ています。膀胱の底の下にあり、尿道の最初の部分を覆っています。 思春期前は筋線維で構成されており、思春期には腺組織が形成され、精子の重要な構成要素である秘密を分泌し、精子の形成を刺激します。 筋線維は括約筋(パルプ)を形成し、性的に興奮すると尿道を圧迫し、射精中に尿が流れるのを防ぎます。 そのため、精液と尿が混ざりません。 生理学男性女性生殖
クーパー腺 2つのエンドウ豆サイズの腺です。 それらは陰茎の根元に位置し、尿道の壁を尿の刺激から保護する粘性のある液体を分泌します。
精子(ギリシャ語から。sperma-種子)は、男性の性腺によって生成され、精子、付属器の液体、および前立腺の分泌物で構成されています。 白っぽいゼラチン状の塊、アルカリ性反応、特有の臭いがあります。 性交中、3〜5mlの精子が3〜5億個の精子を含む膣に排出されます。
精子機能:精子の生存能力と可動性、女性の生殖細胞の受精を確保します。
二次的な男性の性的特徴。これらには、顔の発毛、脇の下と恥骨、喉頭軟骨の成長、および声の変化が含まれます。 これらの兆候の出現は 思春期男の子。 最初に、髪は成長します 上唇、恥骨と脇の下に。 一部の若い男性では、胸骨、肩甲骨、および体の他の部分の領域で髪が成長します。 男性の発毛の有病率と重症度には、個人的および国家的な特徴があり、ある程度、セクシュアリティを反映しています。 巻き毛の短い陰毛はダイヤモンドで成長し、その上部はへそに向けられます(男性タイプの髪の成長)。 喉頭の甲状軟骨はサイズが大きくなり、首の前面に突き出ています(「喉頭隆起」)。 喉頭の軟骨と靭帯の成長は、声の音色の変化(「声の破壊」)を引き起こし、それは低くなります。
二次男性の性的特徴の機能-男性の性別と思春期の指標。
1.2女性の生殖器系の解剖学と生理学
婦人向け生殖器系は、外性器と内性器によって形成され、一次および二次の女性の特徴によって特徴付けられます。
外部の女性生殖器大陰唇、小陰唇、陰核、処女膜、バルトリン腺、乳腺を形成します。
大きな陰唇脂肪を含む皮膚の2つのひだです。 上では、それらは細い巻き毛で覆われた恥骨に入り、下では結合して、膣の後交連を形成します。 膣の後交連と肛門(肛門)の間の空間は会陰と呼ばれます。
大陰唇の間のスリット形成は、生殖器スリットと呼ばれます。 未経産の女性では、大陰唇は閉じており、出産した女性では、いくらか分岐して小陰唇を開きます。 大陰唇の機能:外部の有害な要因の有害な影響から小陰唇を保護し、空気、水、ほこりが膣に侵入するのを防ぎます。 セクシー。
小さな陰唇大陰唇の真ん中に位置し、通常はそれらの間に完全に隠されています。 それらは皮膚の2つの縦方向のひだであり、外観は粘膜に似ています。 小陰唇は外部の刺激に非常に敏感です。 それらの厚さには、結合組織と筋肉組織、血管、感覚神経の末端、および腺の繊維があります。 上の小陰唇はクリトリスを覆い、下の小陰唇は大陰唇の内面と融合します。 小陰唇の間のスリットは呼ばれます 前庭..。 前庭腺の尿道、膣および管はそれに開いています。 小陰唇の機能:保護的でセクシー。 小陰唇は膣への入り口を覆い、水、ほこり、空気が膣に侵入するのを防ぎます。 性的興奮により、血液が充満するために厚くなり、性感帯の感受性が高まります。 陰茎が膣に挿入されると、小陰唇が陰茎を覆い、性感帯の刺激、性的興奮およびオルガスムの増加に寄与します。
クリトリス(緯度から-クリトリス)-生殖器スリットの上隅にある円錐形の形成。 その構造では、クリトリスは男性の生殖器に似ています。 彼の成長は25歳で終わります。 静止状態では、クリトリスの長さと厚さは通常、数ミリメートル以内で変動します。 性的興奮により、クリトリスは密になり、血液の充満によりそのサイズは数倍になります。 陰核よりも陰核の方が3〜4倍敏感な神経終末があります。
クリトリス機能:クリトリスには性的機能があります。 女性の50〜60%では、主な性感帯は陰核にあります。
処女膜(ラテン語-hymen femininusから)小陰唇と膣の境界に位置し、膣の前庭の底を表しています。 処女膜は、膣粘膜のひだによって形成され、多数の弾性線維、血管、神経終末を伴う疎性結合組織で構成されています。 1つまたは複数の穴のある処女膜には約20種類あります。 最初の性交中に、処女膜は中程度の痛みとわずかな出血で破裂(花粉除去)します。 処女膜の機能はほとんど理解されていません。 少女の処女膜はバリア機能を果たし、病原性微生物、空気、ほこり、水が膣に侵入するのを防ぐと考えられています。 思春期後、このバリア機能は大陰唇と小陰唇によって実行され、膣への入り口を覆います。
バルトリン腺形は楕円形で、膣の両側に1つずつあります。 彼らの穴は間の溝にあります 処女膜そして小陰唇の根。
Ba関数NSトリニウム腺:性的興奮により、女性は膣の前庭に潤いを与える粘液を分泌します。 これは、陰茎の膣への自由で痛みのない挿入を促進します。
女性の内臓卵巣、卵管、子宮、膣によって形成されます。 これらの臓器は小さな骨盤にあります。
卵巣(ラテン語の卵巣から)、または女性の性腺は、子宮の左右の小さな骨盤にある対の器官です。 それらは2.5x 1.5 x 1.0 cmの楕円形をしています。胚の卵巣は腹腔内で発達し、その後徐々に骨盤腔内に下降し、女性の生涯を通じて骨盤腔内に留まります。 思春期が始まると、少女の卵巣にグラーフの小胞が形成され、そこで女性の生殖細胞(卵子または卵子)が成長して成熟します。 同時に、1つまたは複数の卵胞が1つまたは2つの卵巣に現れることがあります。 これは、1人、2人、またはそれ以上の双子の子供が生まれたことを説明しています。 2つの独立した卵母細胞から生まれた子供は、3つの卵母細胞(3つの卵母細胞など)から生まれた二卵性双生児と呼ばれます。 同じ卵子から生まれた双子は一卵性双生児と呼ばれ、身体的、生化学的、精神的、その他の指標が非常に似ています。
卵巣機能:女性の生殖細胞の形成と発達; 成長と発達を確実にする2種類の女性ホルモン(エストロゲン、プロゲステロン)の合成と分泌 女性の体; 少量の男性ホルモン(テストステロン)の合成と分泌。これは女性の性的興奮(性欲)を引き起こします。 破裂した卵胞の代わりに、黄体と呼ばれる新しい性腺が形成されます。 それは妊娠の維持と発達を確実にするホルモンを分泌します。 妊娠が起こらなければ、黄体が溶解し、その場所に瘢痕が形成されます。
子宮または卵管-ペアの臓器。 子宮底の角の左右に移動します。 長さは10〜12cm、直径は約2〜3mmです。 卵管の外端は、卵巣と接触している多数のフリンジを備えた漏斗の形をしています。 卵管の壁は、漿液性、筋肉性、粘液性の3つの膜で構成されています。 粘膜は円筒形の繊毛上皮で覆われており、その繊毛は子宮に向かって振動します。 卵管の内腔を通る女性の腹腔。、子宮腔、子宮頸管および膣は外部環境と連絡している。
卵管の機能:上皮の繊毛の振動と卵管の筋線維の収縮により、フリンジに捕らえられた卵子は、腹腔から子宮と精子細胞に移動します。尾、子宮から卵管に移動し、 腹腔..。 原則として、卵管では、男性と女性の生殖細胞の融合が起こり、接合子を形成します(受精)。
子宮洋ナシの形をしており、前の膀胱と後ろの直腸の間の小さな骨盤にあります。 その長さは6〜9cmです。子宮では、底、体、首が区別されます。 子宮頸部は膣の上部に突き出ており、子宮頸管または子宮頸管と呼ばれる管があります。 子宮頸管の一方の端は子宮腔に通じており、もう一方の端は膣に通じています。 子宮頸管は粘液で満たされているため、感染が子宮腔に入るのを防ぎます。 子宮腔は、子宮の底が子宮の底にある三角形の形をしています。 子宮の基部の各隅には、卵管の口があります。 子宮の壁には、外側、中央、内側の3つの層があります。 外層は腹膜カバー、中央によって形成されます 子宮筋層-縦方向および環状配置の平滑筋線維。 妊娠中は子宮の筋層が増加し、出産時に胎児と胎盤を排出するための大きな力が発生します。 出産後、子宮の筋層は元の状態に戻ります。 子宮の内層 子宮内膜(粘膜)卵巣ホルモンの影響下で周期的に変化し、月経周期の終わりに拒絶され、それは小さな血管の露出と子宮(生理学的)出血につながります 月経. 子宮の機能:接合子の粘膜への付着; 胎盤、胚および胎児の成長および発達; 胎児の膜、羊水の形成; 出産と胎盤、月経。
膣(ラテン語-膣から、ギリシャ語-コルポスから)は、長さ7〜13 cm、幅2.5〜4.5 cmの拡張可能なチューブです。出産した女性は、出産していない女性よりも膣の幅が広くなります。 膣には、結合組織、筋肉、粘膜の3つの膜があります。 膣の粘膜は重層扁平上皮で覆われており、腺はありません。 膣は周囲の血管やリンパ管からの発汗液によって保湿されます。 膣の壁は簡単に圧縮および伸ばすことができるため、陰茎の長さと厚さに適応でき、赤ちゃんと胎盤の誕生時にも伸びます。 膣の上端は子宮頸部を覆い、下端は生殖器のスリットに通じています。 子宮頸部の周りには、前部、後部、左、右の4つの膣円蓋があります。 膣円蓋の後部はより深く、精子はその中に蓄積します。 膣の前には膀胱があり、直腸の後ろにあります。
膣機能:保護、導電性、セクシー。 膣の保護機能は、健康な女性の膣に乳酸を分泌する膣棒(微生物)が含まれているという事実によるものです。 したがって、膣の秘密は酸性反応を持っています。 乳酸は、膣に入った病原性微生物の増殖を抑制し、セルフクリーニングプロセスを保証します。 原則として、健康な女性は、口の中よりも膣内の病原性微生物が少ないです。 女性ホルモンの合成に違反して、膣スティックの含有量が減少し、膣分泌物がアルカリ性になり、病原性微生物の発生と膣粘膜の炎症を引き起こします。 膣の酸性環境は、子宮頸部の中性またはアルカリ性環境への精子の移動を確実にします。 膣を通して、子宮頸部と子宮腔、卵子と月経血の分泌物が外部環境に分泌されます。 膣から赤ちゃんと胎盤が生まれ、羊水が放出されます。 性的に成熟した女性では、膣は性的機能を持っています。
二次的な女性の性的特徴。これらには、陰毛と脇毛の成長、皮膚の下の特定の種類の脂肪沈着、幅の骨盤の成長、乳腺の成長、および月経機能の確立が含まれます。 髪の伸び。 皮下脂肪層。 骨盤の骨。 14歳までに、少女の陰毛は、短くて粗い巻き毛と脇の下のまっすぐな髪で成長します。 恥骨では、髪の毛は三角形の形で成長し、その基部には水平線があります(女性タイプの髪の成長)。 皮膚の下、特に骨盤領域に脂肪組織が沈着し、骨盤の骨が水平方向に拡張することで、少女の体は丸みを帯びた形になり、女性の体型を形成します。 乳腺(Lat。-mammaeから)汗腺の派生物ですが、機能的には生殖器に関連しています。 人は乳腺の1つのペアを持っています 胸したがって、それらは乳腺とも呼ばれます。 少女と少年の出生時までに、各乳腺の直径は0.4〜2.5 cmです。男性の場合、乳腺は一生基本的な状態のままです。 少女では、乳腺は視床下部、下垂体、卵巣、副腎、甲状腺のホルモンの影響下で10〜12歳で発達し始めます。 月経が始まると、乳腺の成長が加速します。 乳腺は妊娠の終わりまでに最大の発達に達します。 授乳が終わると、乳腺のサイズが小さくなります。 腺の前面には乳首があり、その頂点には乳管の出口開口部があります。 乳首は、乳頭円または乳輪と呼ばれる色素沈着した皮膚のパッチに囲まれています。 乳輪の皮膚はでこぼこしています。これは、皮脂腺とその開口部が埋め込まれていることが原因です。 乳輪と乳首の皮膚には、神経終末と平滑筋線維が含まれています。 筋線維の収縮に伴い、乳首は密になり、長さが長くなります。 これにより、授乳中に赤ちゃんが乳房を吸いやすくなります。 乳腺の腺組織は小葉で構成されており、その排泄管は乳頭の頂点で開く乳管に接続されています。 通常、乳首には8〜10個の乳管出口があります。 乳腺には、形や大きさの点で個々の特徴があります。 彼らは性感帯を発達させました。
女性の乳腺の機能:分泌、美的および性的。 分泌機能乳腺は、妊娠の終わりと出産後に現れ、初乳と乳汁の分泌で構成されます。 初乳と乳汁の形成と排泄のプロセスは、授乳と呼ばれます。 初乳は、アルカリ性反応の濃い黄色がかった液体です。 妊娠末期と出産後数日で分泌されます。 ロストは、生まれたばかりの赤ちゃんにとって欠かすことのできない食べ物です。 母乳と比較して、初乳には多くのタンパク質、ビタミン、抗体、酵素、ミネラルが含まれており、脂肪と炭水化物が少なくなっています。 ミルクはアルカリ反応のある白い液体です。 乳汁の分泌は、出生後2〜3日で始まり、女性が授乳している間、出生後2〜3年間続く可能性があります。 1。5年後、ミルクの栄養価は低下します。 ミルクの分泌とその分離は、中枢によって調節される無条件および条件付き反射です 神経系..。 吸うという行為は、乳首と乳輪の神経線維の末端を刺激します。 それらからの神経インパルスは大脳皮質に行き、そこから視床下部と下垂体に行き、そこでミルクの分泌(プロラクチン)とミルクダクトへのミルクの放出(オキシトシン)に関与するホルモンが生成されます。 否定的な感情は減少し、肯定的な感情は乳汁分泌を増加させます。 月経(ラテン語のmenstruusから-毎月)-思春期に達した少女と出産可能年齢の女性の子宮から膣を介した定期的な血液の排出。 月経は、卵巣から腹腔への女性の生殖細胞の放出(排卵)に関連しています。 月経周期は、前の期間の初日から次の月経の初日までの時間です。 月経と月経周期は個別です。 ほとんどの女性の場合、月経周期は26〜30日ですが、それほど頻繁ではありません-21〜24日(短縮)または30日以上(延長)です。 周期の途中で、卵巣で成熟した卵胞が破裂し、卵子が腹腔内に放出されます。 この段階で妊娠する可能性が最も高くなります。 月経の期間は4-6日で、失われる血液の量は約50mlです。 月経の最初と最後の日に生成される血液は少なくなります。 月経の初日に出血がより顕著になることがあります。 月経の期間と失血量は、さまざまな要因(一般的および婦人科の病気、否定的な感情など)の影響下で変化する可能性があります。 女の子の最初の期間は初潮と呼ばれます。 ほとんどの女の子は何らかの形を経験します 不快感これは、体内の生理学的プロセスだけでなく、この新しい現象の認識と評価にも起因します。 月経の心理的な準備ができている女の子は、月経が正常であると認識しており、新しい有望な成人生活に入っていることを示しています。 原則として、健康な女性は月経によく耐えます。 しかし、月経の初日、特に若い女の子では、下腹部にわずかな倦怠感、脱力感、痛みがあるかもしれません。 月経前に乳腺が痛む可能性があります。 月経中の一部の女性は、より感情的で、気難しいようになり、ささいなことに動揺する可能性があります。 しかし、これらは病気の症状ではありません。 したがって、あなたは通常の生活、仕事、そして休息を送る必要があります。 ただし、月経中は、身体的ストレスの増加(ウェイトリフティング、ジャンプ、サイクリング、乗馬など)を避ける必要があります。泳いだり、入浴したり、辛い食べ物を食べたりすることはできません。 どうぞ 薬子宮の筋肉のけいれんによる痛みを軽減します(shpaなしなど)。 すべての月経中の女性は、彼女の期間の期間と月経周期とそれらの特徴を知っている必要があります。 これを行うには、ポケットカレンダーで生理の最初と最後の日をマークする必要があります。 排卵に伴う月経周期の途中で、膣からわずかに血液が排出されることがあります。
Allbest.ruに投稿
同様の文書
全般的 解剖学的構造:精巣、精巣上体、精子管、副性腺、陰茎(陰茎)および包皮。 精子形成の生理学的現象。 男性の反芻動物と片足の動物の生殖器の構造の特徴。
テスト、2009年5月5日追加
男性と女性の生殖器。 生殖システムは、生殖製品の生産に関与し、受精のプロセスを確実にし、人間の生殖に寄与する器官とシステムの複合体です。 人間の性的二形の程度。
2010年2月19日にプレゼンテーションが追加されました
女性の生殖器の一般的な特徴、子宮とその付属肢の構造と機能。 粘膜と筋肉膜の特徴。 子宮と腹膜およびその靭帯装置との関係。 血流、リンパの流れ、臓器の神経支配。 卵巣の構造と機能。
要約、2011年9月4日追加
泌尿生殖器系への統合、発達と位置の観点からの泌尿生殖器と生殖器の相互のコミュニケーション。 腎臓の構造の特徴、それらの構造的および機能的単位としてのネフロン。 膀胱、男性と女性の生殖器の構造。
2017年5月22日にプレゼンテーションが追加されました
科学としての解剖学と生理学。 内部環境の役割、神経質で 循環系細胞の必要性を生物全体の必要性に変換することにおいて。 体の機能システム、それらの調節および自己調節。 人体の一部、体腔。
プレゼンテーションは2015年9月25日に追加されました
男性生殖器の内部構造:前立腺、陰嚢、陰茎。 女性の内臓の構造。 会陰から血液を運ぶ静脈。 聴覚器官の機能。 人間の発達過程における聴覚。
要約、2013年10月16日追加
人間の腎臓の解剖学と形態。 生理学と機能。 腎臓は一種の内分泌腺のようなものです。 体からの代謝最終産物の除去。 水分バランス、酸塩基状態、血圧レベルの調節。
タームペーパー追加2009年8月8日
各年齢段階での身体の生理学的機能の発達。 主題としての解剖学と生理学。 人体とその構成構造。 代謝とエネルギーとその 年齢の特徴..。 身体機能のホルモン調節。
チュートリアル、2010年12月20日追加
動物の精子の多様性。 精子の主な機能。 男性の生殖細胞の形成。 人間の精子、それらの構造、機能、動き、平均余命。 植物界の精子。 生殖細胞の発達のスキーム。
要約追加2013年9月18日
細胞の構造の説明、および生物で使用されるいくつかの有機化合物。 人類の生理学と解剖学、多くの重要な器官の機能の特徴。 体内の相互作用と代謝。 水生生物環境。
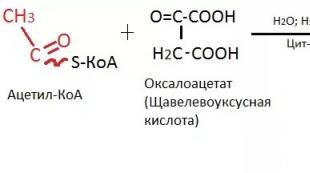
女性の生殖機能 卵巣では熟すので、主に卵巣と子宮の活動のために実行されます 卵、そして子宮では、卵巣から分泌されるホルモンの影響下で、知覚に備えて変化が起こります 受精卵. 生殖期間子孫を再生する女性の体の能力によって特徴付けられます。 この期間の期間は17-18から45-50年です。 生殖または出産の期間の前に、女性の人生の次の段階があります。 子宮内; 新生児(最長1年); 子供時代(8-10歳まで); 思春期前と 思春期年齢(17〜18歳まで)。 生殖期間が始まります 更年期、を区別する 閉経前, 閉経と 閉経後.
月経周期 -女性の体の複雑な生物学的プロセスの症状の1つ。 月経周期は、生殖器系のすべての部分の周期的な変化によって特徴付けられ、その外部症状は 月経.
月経 - これは 生殖管からの血の分泌物二相性月経周期の終わりに子宮内膜の機能層の拒絶の結果として定期的に発生する女性。 最初の月経( 初潮)10〜12歳で観察されますが、この後1〜1。5年以内に、月経が不規則になり、その後規則的になることがあります 月経周期.
月経の初日従来、月経周期の初日に服用していました。 したがって、サイクルの期間は、次の2つの月経の最初の日の間の時間です。 女性の60%、平均 月経周期の持続時間は28日で、21日から35日まで変動します。 月経日の失血量 40〜60 ml、平均50ml。 通常の月経の期間 2日から7日。
卵巣。月経周期の間に卵巣で成長が起こります 卵胞と 卵の成熟、結果として準備が整います 受精..。 同時に、性ホルモンは卵巣で生成され、受精卵を受け取ることができる子宮の内壁に変化をもたらします。
性ホルモン (エストロゲン, プロゲステロン, アンドロゲン) それは ステロイド、彼らの教育には 幻の細胞濾胞、内層と外層の細胞。 性ホルモン卵巣によって合成されたものは、標的の組織や臓器に影響を及ぼします。 これらには以下が含まれます 性器、 初めに 子宮, 乳腺, 海綿骨, 脳, 内皮と 血管平滑筋細胞, 心筋, レザーと彼女 付属物(毛包および皮脂腺)など。標的細胞へのホルモンの直接接触および特異的結合は、対応する受容体との相互作用の結果です。
生物学的効果は、遊離(非結合)画分によって与えられます エストラジオールと テストステロン(1%)。 卵巣ホルモンの大部分(99%)が結合しています。 輸送は特別なタンパク質によって行われます- ステロイド結合グロブリンおよび非特定の輸送システム- アルブミンと 赤血球.
描く: 優勢な濾胞の発達段階。
NS - 原始濾胞; NS - 前胞状卵胞; v- 胞状卵胞; NS - 排卵前の卵胞: 1 - 卵母細胞, 2 - 顆粒膜細胞 (粒状ゾーン), 3 - 卵胞膜細胞, 4 - 基底膜.
エストロゲンホルモン に貢献する 生殖器の形成、 発達 第二次性徴思春期の間に。 アンドロゲン外観に影響を与える 恥骨と脇の下の髪. プロゲステロン 月経周期の分泌期を制御し、着床のために子宮内膜を準備します。 性ホルモンは妊娠と出産の発達に重要な役割を果たします。
卵巣の周期的な変化には、3つの主要なプロセスが含まれます:
1. 卵胞の成長と優勢な卵胞の形成.
- 排卵.
- 黄体の形成、発達および退行。
少女の誕生時には、卵巣には200万個の卵胞があり、その99%が卵胞にさらされています。 閉鎖一生を通じて。 閉鎖のプロセスは、その発達の段階の1つで卵胞の逆発達として理解されています。 やがて 初潮卵巣には約20万から40万の卵胞があり、そのうち300から400は排卵の段階まで成熟します。
卵胞発達の以下の主要な段階を区別するのが通例です: 原始濾胞, 前胞状卵胞, 胞状卵胞, 排卵前の卵胞.
原始濾胞 l卵胞および顆粒(顆粒)上皮に位置する未熟卵で構成されています。 外では、卵胞は結合膜に囲まれています( 卵胞膜細胞)。 各月経周期の間に、3〜30個の原始卵胞が成長して形成され始めます 前洞、 また 主要な、卵胞。
前胞状卵胞. 成長の始まりとともに 原始濾胞前胞状段階に進み、そして 卵母細胞拡大し、と呼ばれる膜に囲まれています 光沢のあるシェル (透明帯)。 顆粒膜上皮の細胞は増殖し、周囲の間質から卵胞膜が形成されます。 この成長は、エストロゲン産生の増加を特徴としています。 前胞状卵胞の顆粒膜層の細胞は合成することができます ステロイドエストロゲンはアンドロゲンやプロゲステロンよりもはるかに多く合成されますが、3つのクラスがあります。
アントラル、 また 二次、卵胞 ..。 それはさらなる成長によって特徴付けられます:顆粒膜層の細胞の数、生成 卵胞液..。 卵胞液は顆粒膜層の細胞間空間に蓄積し、空洞を形成します。 卵胞形成のこの期間(月経周期の8-9日目)の間に、性ステロイドホルモン、エストロゲンおよびアンドロゲンの合成が注目されます。
性ホルモンの合成に関する現代の理論によれば、アンドロゲンは卵胞膜細胞で合成されます- アンドロステンジオンと テストステロン..。 次に、アンドロゲンは顆粒膜層の細胞に入り、それらの中でそれらはエストロゲンに芳香化されます。
優勢な濾胞 ..。 原則として、そのような卵胞の1つは、多くの胞状卵胞から形成されます(サイクルの8日目までに)。 それは最も大きく、顆粒膜層の細胞とFSH、LHの受容体の最大数を含んでいます。 優勢な卵胞は、血管が豊富な卵胞膜を持っています。 卵巣における優勢な排卵前卵胞の成長と発達とともに、残りの(90%)成長中の卵胞の閉鎖のプロセスが並行して起こります。
月経周期の最初の日の優勢な濾胞は2mmの直径を持っています、そしてそれは排卵の時までに14日以内に平均21mmに増加します。 この間、卵胞液の量が100倍に増加します。 それはエストラジオールとFSHの含有量を急激に増加させ、成長因子も決定されます。
排卵 - 卵胞前優性の破裂(三次)卵胞とそれからの卵子の放出。 排卵の時までに、卵母細胞でプロセスが起こります 減数分裂. 排卵は出血を伴う卵胞膜を取り巻く破壊された毛細血管から。 排卵は、エストラジオールの排卵前のピークが形成されてから24〜36時間後に発生すると考えられています。 排卵前濾胞の壁の菲薄化と破裂は、酵素の影響下で起こります コラゲナーゼ..。 また、役割を果たします プロスタグランジンF2a卵胞液に含まれるEr。 顆粒膜細胞で産生されるタンパク質分解酵素; オキシトシンと リラキシン.
卵子の放出後、結果として生じる毛細血管はすぐに卵胞腔に成長します。 顆粒細胞が露出している 黄体化:それらは細胞質の体積を増加させ、形成します 脂質封入体..。 LHは、顆粒膜細胞のタンパク質受容体と相互作用し、黄体形成のプロセスを刺激します。 このプロセスは形成につながります 黄体.
黄体 - 一過性内分泌腺月経周期の長さに関係なく、14日間機能します。 妊娠していない場合、黄体は退行します.
したがって、主な女性の性ステロイドホルモンは卵巣で合成されます- エストラジオールと プロゲステロン、 と アンドロゲン.
V 月経周期のIフェーズ、月経の初日から排卵の瞬間まで続く、体はエストロゲンの影響下にあり、II(排卵から月経の開始まで)ではエストロゲンに加わります プロゲステロン黄体の細胞から分泌されます。 月経周期の最初の段階は、 卵胞、 また 卵胞、サイクルの第2フェーズは 黄体.
月経周期の間、末梢血中のエストラジオールの含有量には2つのピークがあります。1つ目は顕著な排卵前周期であり、2つ目はそれほど顕著ではなく、月経周期の第2段階の中間です。 サイクルの第2段階で排卵した後、プロゲステロンが主なものであり、その最大量は排卵後4〜7日目に合成されます。
卵巣でのホルモンの周期的な分泌は、子宮の内層の変化を決定します。
子宮内膜(子宮内膜)の周期的変化. 子宮内膜は次の層で構成されています。
基底層 それは月経中に拒絶されません。 月経周期中の細胞から、子宮内膜の層が形成されます。- 表面層子宮腔を裏打ちするコンパクトな上皮細胞で構成されています。
- 中級、 また 海綿状の層.
最後の2つの層は機能層を構成します。機能層は月経周期の間に大きな周期的変化を受け、月経中に拒絶されます。
月経周期のフェーズIでは、子宮内膜は腺と間質の薄い層です。 サイクル中の子宮内膜の変化には、次の主要な段階があります:
1) 増殖期;
2) 分泌期;
3) 月経.
増殖期 ..。 成長中の卵胞によるエストラジオールの分泌が増加すると、子宮内膜は増殖性の変化を起こします。 基底層の細胞は活発に増殖します。 細長い管状腺を備えた新しい表面の緩い層が形成されます。 この層はすぐに4〜5倍厚くなります。 円柱上皮で裏打ちされた管状腺は細長くなっています。
分泌段階 ..。 卵巣周期の黄体期では、プロゲステロンの影響下で、腺の屈曲度が増加し、その内腔が徐々に拡大します。 間質の細胞は、体積が増加し、互いに接近します。 腺の分泌が増加します。 腺の内腔には大量の分泌物が見られます。 分泌の強さに応じて、腺は強く複雑なままであるか、鋸歯状になります。 間質の血管新生が増加しています。 分泌の初期、中期、後期を区別します。
月経 ..。 それ 子宮内膜の機能層の拒絶..。 月経の開始とプロセスの根底にある微妙なメカニズムは不明です。 月経の開始の内分泌基盤は、黄体の退行によるプロゲステロンおよびエストラジオールレベルの顕著な減少であることが見出された。
月経に関与する主な局所メカニズムは次のとおりです:
1) らせん状動脈の色調の変化;
2) 子宮内の止血のメカニズムの変化;
3) 子宮内膜細胞のリソソーム機能の変化;
4) 子宮内膜の再生.
月経の開始に先立って、らせん状細動脈が激しく狭くなり、虚血および 落屑子宮内膜。
月経周期の間に、内容は変わります リソソーム子宮内膜の細胞で。 リソソーム酵素が含まれており、その一部はプロスタグランジンの合成に関与しています。 プロゲステロンレベルの低下に応じて、これらの酵素の放出が増加します。
子宮内膜の再生月経の最初から観察されました。 月経の24時間目の終わりまでに、子宮内膜の機能層の2/3が拒絶されます。 基底層子宮内膜の再生の基礎となる間質の上皮細胞が含まれています。これは通常、周期の5日目までに完全に完了します。 並行して完了する 血管新生破れた細動脈、静脈、毛細血管の完全性が回復します。
卵巣と子宮の変化は、月経機能を調節するシステムの二相活動の影響下で発生します。 大脳皮質, 視床下部, 下垂体..。 したがって、女性の生殖システムには5つの主要なリンクがあります。 大脳皮質, 視床下部, 下垂体, 卵巣, 子宮..。 生殖器系のすべてのリンクの相互関係は、性ホルモンと性腺刺激ホルモンの両方の受容体の存在によって保証されています。
生殖器系の調節における中枢神経系の役割は長い間知られています。 これはによって証明されました 排卵障害さまざまな急性および慢性のストレスを伴う、 月経周期の違反気候地理的ゾーンを変更するとき、仕事のリズム; よく知られている 戦時中の月経の停止..。 精神的に不安定で赤ちゃんを産むことを切望している女性も月経をやめるかもしれません。
大脳皮質と 視床下部外の脳構造(大脳辺縁系、海馬、扁桃体など)、エストロゲン、プロゲステロン、アンドロゲンの特定の受容体が同定されています。 合成、排泄、代謝はこれらの構造で起こります。 ニューロペプチド, 神経伝達物質そしてそれらの受容体は、合成と分泌に選択的に影響を及ぼします 視床下部放出ホルモン.
性ステロイドと組み合わせて、それらは機能します 彼女rotransmitters :ノルエピネフリン, ドーパミン, ガンマアミノ酪酸, アセチルコリン, セロトニンと メラトニン. ノルエピネフリン 放出を刺激する ゴナドトロピン放出ホルモン (GTRG)前核視床下部のニューロンから。 ドーパミンと セロトニン頻度を減らし、生産の振幅を減らします GTRG月経周期のさまざまな段階で。
ニューロペプチド(内因性オピオイドペプチド, ニューロペプチドY、コルチコトロピン放出因子およびガラニン)も生殖器系の機能に影響を及ぼし、したがって視床下部の機能に影響を及ぼします。 内因性オピオイドペプチド 3種類( エンドルフィン, エンケファリンと ダイノルフィン)脳内のアヘン剤受容体に結合することができます。 内因性オピオイドペプチド( イメージインテンシファイア)コンテンツに対する性ホルモンの影響を調整する GTRGフィードバックメカニズムにより、下垂体、特に下垂体による性腺刺激ホルモンの分泌をブロックします LH、視床下部におけるGTRHの分泌を遮断することによって。
インタラクション 神経伝達物質と ニューロペプチド生殖年齢の女性の体内で定期的な排卵周期を提供し、視床下部によるGTRHの合成と分泌に影響を与えます。
視床下部には、刺激を分泌するペプチド作動性神経細胞が含まれています( リベリン)およびブロッキング( スタチン)神経ホルモン- 神経分泌..。 これらの細胞はニューロンと内分泌細胞の両方の特性を持っており、血流から来る信号(ホルモン)と脳内の神経伝達物質と神経ペプチドの両方に反応します。 神経ホルモンはニューロンの細胞質のリボソームで合成され、軸索に沿って末端に輸送されます。
ゴナドトロピン放出ホルモン( リベリン)は、FSHとLHが合成される下垂体の性腺刺激ホルモン機能を調節する神経ホルモンです。 放出ホルモンLH( ルリベリン)が強調表示され、合成され、詳細に説明されています。 卵胞刺激ホルモン放出ホルモンを分離して合成する、または フォリーベリン、これまでのところ失敗しました。
ゴナドリベリンの分泌には脈動する特徴があります。数分間続くホルモンの分泌増加のピークは、比較的低い分泌活性の1〜3時間間隔で置き換えられます。 GnRH分泌の頻度と振幅は、エストロゲンのレベルによって調節されます。
下垂体前葉によるプロラクチンの分泌を制御する神経ホルモンは、 プロラクチン阻害ホルモン(係数)、または ドーパミン.
生殖器系の重要なリンクは下垂体の前葉です- 下垂体前葉、性腺刺激ホルモンが分泌される、 卵胞刺激ホルモン (FSH, フォリトロピン) 黄体形成ホルモン (LH, ルトロピン) と プロラクチン (Prl)、卵巣と乳腺の機能を調節します。 3つのホルモンはすべてタンパク質物質です( ポリペプチド)。 性腺刺激ホルモンの標的腺は卵巣です。
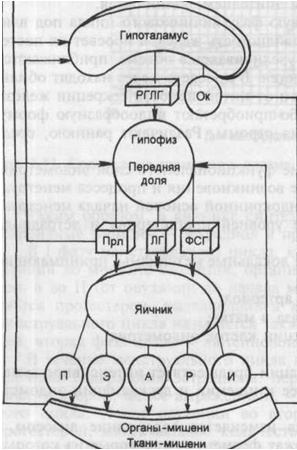 描く: 生殖器系の機能(図)。
描く: 生殖器系の機能(図)。
WGLG - ホルモンの放出; わかった - オキシトシン; Prl- プロラクチン; FSH - 卵胞刺激ホルモン; NS - プロゲステロン; NS - エストロゲン; NS- アンドロゲン; NS - リラキシン; と - インヒビン; LH - 黄体形成ホルモン.
下垂体の前葉では、それらも合成されます サイロトロピック(TSH) と 副腎皮質刺激ホルモン(ACTH)ホルモン、および成長ホルモン。
FSHは、卵胞の成長と成熟を刺激し、卵胞顆粒膜細胞の表面でのFSHおよびLH受容体の形成を促進し、成熟卵胞のアロマターゼの含有量を増加させ、芳香化プロセスを刺激することにより、アンドロゲンのエストロゲンへの変換を促進します。卵胞の成長において抑制的かつ刺激的な役割を果たすインヒビン、アクチビン、およびインスリン様成長因子-1の産生を刺激します。
LHは刺激します:
卵胞膜細胞におけるアンドロゲンの形成;
FSHによる排卵;
黄体形成中の顆粒膜細胞のリモデリング;
黄体におけるプロゲステロンの合成.
プロラクチン 乳腺の成長を刺激します授乳は、黄体のLH受容体の形成を活性化することにより、黄体によるプロゲステロンの分泌を制御します。
下垂体前葉によるプロラクチンの合成は、ドーパミンの強壮性遮断制御下にあります、または プロラクチン阻害因子..。 プロラクチン合成の阻害は、妊娠中、授乳中に停止します。 プロラクチン合成の主な刺激因子は、視床下部で合成されるチロリベリンです。
視床下部-下垂体系と卵巣の周期的変化は相互に関連しており、フィードバックの種類に応じてモデル化されています。
次の種類のフィードバックが区別されます:
1) "長いループ「フィードバック-卵巣のホルモンと視床下部の核の間;卵巣のホルモンと下垂体の間;
2)"短いループ"-下垂体前葉と視床下部の間;
3)"超短ループ"-GTRHと視床下部の神経細胞の間。
これらすべての構造の関係は、それらの中にある性ホルモン受容体の存在によって決定されます。
生殖年齢の女性は、卵巣と視床下部-下垂体系の間に負と正の両方のフィードバックがあります。 負帰還の例は 下垂体前葉によるLHの放出の増加サイクルの初期卵胞期における低エストラジオールレベルに応答して。 正のフィードバックの例は LHサージ排卵の最大値に応じて 血中のエストラジオールの含有量.
生殖器系の状態は、機能診断テストの評価によって判断することができます。 基礎体温, 瞳孔症状と カリオピクノティックインデックス.
基礎体温 朝の直腸で測定ベッドから出る前に。 排卵月経周期では、基礎体温は周期の黄体期に0.4〜0.6°C上昇し、第2期全体を通して持続します(図を参照)。 月経の当日またはその前日には、基礎体温が低下します。 妊娠中の基礎体温の上昇は、プロゲステロンの影響下での視床下部の体温調節中心の興奮によって説明されます。
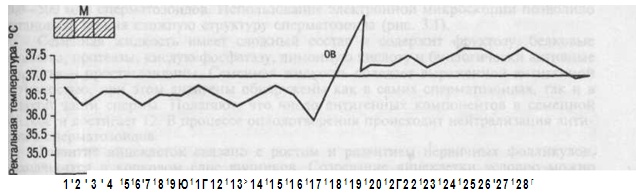 描く: 二相性サイクルにおける直腸温。 M-月経; OV-排卵。
描く: 二相性サイクルにおける直腸温。 M-月経; OV-排卵。
瞳孔症状 子宮頸管の粘液の変化を反映しています。 エストロゲンの影響下で、透明な硝子体粘液が子宮頸部に蓄積し、それが子宮頸部の外部開口部の拡張を引き起こします。 粘液の最大量は、周期の排卵前の日に観察され、外側の開口部は暗くなり、瞳孔に似ています。 サイクルの第2段階では、プロゲステロンの影響下で、粘液の量が減少するか、完全に消失します。 粘液はゴツゴツした構造になっています。 瞳孔の症状には3つの程度があります:+、++、+++.
Karyopyknoticインデックス ..。 卵巣ホルモンの影響下で、周期的な変化は膣粘膜、特にその上部3分の1でも発生します。 V 膣スミア、以下が発生する可能性があります 扁平上皮細胞の種類 : NS) 角質化、 NS) 中級、v) 基礎、 また 萎縮性..。 最初のタイプの細胞は、卵巣によるエストロゲンの分泌が増加するにつれて優勢になり始めます。 細胞要素の定量的比率の決定に基づいて、エストロゲンホルモンによる体の飽和度またはそれらの不足を判断することができます。 角質化細胞の最大数は、排卵前の日に検出されます-80-88%、増殖の初期段階-20-40%、分泌の後期-20-25%。
情報は不完全ですか? それを試してみてください Google検索 .