Герпесвирусные инфекции. Размножение вируса простого герпеса
Вирус - особая форма жизни, которая не имеет собственного аппарата для размножения, поэтому вирусные частицы проникают в клетку - хозяина и используют ее возможности и аппарат для собственного размножения.
Вне клетки вирус можно сравнить с магнитофонной кассетой, лежащей на подоконнике: на ленте записана какая-то информация, но музыки не услышишь. Пока кассета лежит на подоконнике, с ней ничего не происходит. Аудиокассета не может самостоятельно скопироваться или заиграть, для этого нужен магнитофон. Так и вирус вне клетки не может размножиться, кроме того, в нем не происходит никаких метаболических процессов: ему не нужно ни дышать, ни пить, не есть.
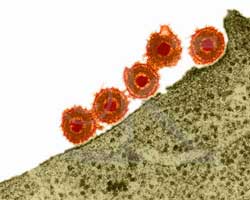
Воспроизводство вируса простого герпеса осуществляется в несколько последовательных стадий, сменяющих друг друга. На первом этапе вирус "приклеивается" (адсорбируется) к клетке - хозяину, затем
проникает внутрь, "сбрасывая свои одежды" (внешние оболочки). Потом проникает в ядро клетки, где и синтезируется новый вирус. На последнем этапе вирус покидает клетку.
После полового акта или орального секса вирусы в большом количестве попадают на слизистые или кожные покровы неинфицированного полового партнера. Множество вирусных частиц соударяются с эпителиальными клетками человека. На первом этапе часть вирусных частиц при помощи "шипиков" гликопротеидов прилипают (адгезируются) к наружной мембране клетки. На 1000 соударений с клеткой лишь в 1 случае вирус прикрепляется к ее наружной оболочке. На этом этапе, который называется "этап обратимой адгезии" еще возможен отрыв вируса от наружной мембраны клетки
Постепенно наружная оболочка вируса сливается с мембраной клетки, изменяя ее поверхностное строение.
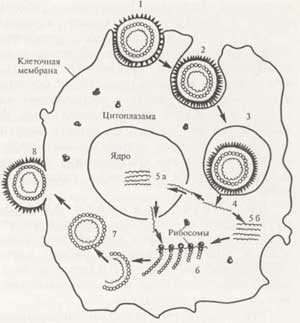
Рисунок
"Стадии репродукции вирусов (схема)".
1 - адсорбция вируса на клетке;
2 - проникновение вируса в клетку;
3 - вирус внутри вакуоли клетки;
4 - "раздевание" вируса;
5 - репликация вирусной нуклеиновой кислоты в ядре
клетки (а)
6 - синтез вирусных белков на рибосомах клетки;
7 - формирование вируса;
8 - выход вируса из клетки путем почкования.
Вирус "раздевается", скидывая не нужные ему внутри клетки защитные оболочки. На вирусной ДНК остается один лишь 20 гранный капсид.
Затем вокруг вируса, впрыснутого в клетку образуется маленький пузырек - вакуоль. В этом пузырьке вирус транспортируется к клеточному ядру. Капсид прикрепляется к ядру клетки. Он растворяет часть оболочки ядра клетки и вирусная ДНК оказывается внутри ядра.
Через 2 часа от момента проникновения вируса в клетку, вирусная ДНК блокирует в ядре клетки производство собственной ДНК и заставляет Клетку производить ДНК вируса.
В течение 7-9 часов ядро Клетки накапливает вирусную ДНК. Здесь же образуется и капсид вируса. Структуры ядра разрушаются. Через 9 часов от начала момента синтеза вирусной ДНК, вирусы выходят из ядра в цитоплазму клетки. Здесь они используя ядерную мембрану и мембрану клетки приобретают внешнюю оболочку - суперкапсид.
Через 24 часа вирусы методом почкования покидают клетку. Из - за повреждений клетка наполняется жидкостью и вскоре гибнет.
Одна клетка дает несколько миллионов вирусов. Из эпителиальной клетки на месте проникновения вирусы попадают в кровяное русло, приклеиваясь к эритроцитам и с кровью разносятся по всему организму.
Второй путь распространения вирусов - по нервным волокнам. По чувствительным веточкам вирус попадает в нерв, где с током аксоплазматической жидкости проникает в чувствительные нейроны ганглиев крестцового сплетения. В нервных клетках вирус сбрасывает себя капсид и суперкапсид и "живет" в клетках в виде двуспиральной молекулы ДНК. В таком состоянии иммунная система не в состоянии обнаружить вирус и удалить его из организма.
В случае действия внешних факторов: алкоголь, стресс, травма, заболевание, приводящее к снижению иммунитета, вирус начинает размножаться в крестцовых ганглиях и с током аксоплазматической жидкости дрейфует в сторону половых органов. Где уже в эпителиальных клетках происходит рецидив инфекции.
Источники: Схема репродукции вирусов позаимствована из учебника Микробиологии Воробьева, Быкова, Пашкова и Рыбаковой М, "Медицина" 1994 год стр. 47
Исследовательский медицинский университет им.
Н.И.Пирогова
Минздравсоцразвития России
КАФЕДРА МИКРОБИОЛОГИИ И ВИРУСОЛОГИИ
Заведующая кафедрой – Кафарская Людмила Ивановна
Реферат на тему: «простой герпес»
Выполнил студент 244 группы
Колпаков Дмитрий
Проверил преподаватель:
Харитонов Анатолий Аркадьевич
Москва 2013
1)Таксономическое положение – семейство род, вид 2)Морфология; 3)Репродукция герпесвирусов 4)Антигенные свойства; 5)Патогенез; 6)Диагностика 7)Лечение 8)Иммунитет
1)Таксономическое положение – семейство род, вид
Простой герпес (herpes simplex), семейство herpesviridae, подсемейство альфа-herpesviridae, род simplexvirus, ГВЧ 1 и 2 типа
2)Морфология;
Крупные оболочечные ДНК содержащие вирусы. Размеры вириона 150-200 нм. Вирус имеeт капсид из 162 капсомеров, а также суперкапсид. Капсид содержит линейную двухнитевую ДНК, в которой присутствуют около 80 генов. Тип симметрии кубический.
Культивируются в культурах живых клеток и вызывают их многоядерность. Чувствительны к нагреванию, быстро инактивируются УФидезинфицирующими средствами.
3)Репродукция герпесвирусов
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко. Ранняя стадия репродукции герпесвирусов. В ранней стадии синтезируются «ранние белки», кодируемые проксимальной третью молекулы ДНК. Они проявляют регуляторные свойства, включая активацию транскрипции других участков вирусного генома, кодирующих ДНК-полимеразу и ДНК-связывающие белки. Поздняя стадия репродукции герпесвирусов. В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов). Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов - формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
4)Антигенные свойства;
Типоспецифические антигены представлены гликопротеинами капсида, на основе которых и выделяют два типа вирусов: HSV-1 и HSV-2 . Группоспецифические антигены представлены нуклеопротеидами. Поверхностный гликопротеин gpC-группоспецифический, поверхностные гликопротеиды gpB, gpD- типоспецифические.
5)Патогенез;
Вирус герпеса не способен проникать через неповреждённый роговой слой кожи, что обусловлено отсутствием на нём специфических рецепторов. Герпес передаётся при прямом контакте повреждённых покровов с поражёнными участками кожи или с биологическими жидкостями инфицированного человека. Вирус HSV-2 типа передаётся исключительно транскутанно (через кожу).
Внешняя оболочка вируса сливается с клеточной мембраной. Далее вирусный нуклеокапсид оказывается в нейроплазме, где и происходит высвобождение вирусной ДНК. Далее она транспортируется по дендритам нервных окончаний в тело чувствительного нейрона, находящееся в сенсорном ганглии, где встраиваются в его генетический аппарат навсегда. После проникновения вируса начинается процесс его активного воспроизводства в клетке - персистенция. При лабиальных поражениях характерна персистенция вируса в нейроцитах чувствительных ганглиев тройничного нерва, а при генитальных - поясничных.
У большинства людей воспроизводство и выделение вируса сразу после инфицирования происходит бессимптомно. Это может произойти более чем за неделю до или после появления первых симптомов в 50 % случаев. Возбудитель интенсивно в нем размножается, запуская литический, продуктивный тип инфекции. Происходит очаговая дегенерация эпителия: клетки увеличиваются в размерах, затем погибают, образуя очаги некроза.
Вирусы герпеса имеют циклические периоды активности (в течение 2-21 дней формируются пузырьки, содержащие вирусные частицы) и периоды ремиссии, во время которых язвочки пропадают. Генитальный герпес (HSV-2-типа), чаще протекает бессимптомно, хотя вирус размножается и может передаваться другим людям. Это наиболее характерно для вируса HSV-2-типа. Рецидивы заболевания не определены во времени, хотя выявлены некоторые триггеры заболевания. К таким факторам относится воздействие иммунодепрессантов (см. ниже ). При рецидиве вирус, находящийся в латентной стадии, активируется, в результате чего образуется множество инфекционных частиц, движущихся по отросткам нейрона, из которых они в дальнейшем переходят в эпителий кожи и слизистых оболочек. Часто снова возникают пузырьки, сопровождающиеся некротическим повреждением эпителия. Цикл репродукции вируса - 10 часов. С течением времени у организма формируется противовирусный иммунитет и частота и тяжесть рецидивов уменьшается.
Носительство чаще всего продолжается в течение первых 12 месяцев после инфицирования. Продолжительность такого носительства при иммунодефицитах (например при ВИЧ-инфекции ) больше. После заражения организм начинает синтезировать антитела против конкретного HSV-типа вируса, предотвращая распространение инфекции. В случае заражения вирусом HSV-1-типа, такая сероконверсия (выработка антител) защитит организм от прочих инфекционных процессов, вызываемых этим вирусом, таких как генитальный герпес, герпетический кератит и панариций.
Антитела, которые вырабатываются после первоначальной герпетической инфекции, предотвращают заражение тем же типом вируса: у людей, перенёсших ротолицевой герпес HSV-1-типа не бывает панариция или генитального герпеса, вызываемого HSV-1.
В моногамном браке, серонегативная женщина имеет риск заражения от серопозитивного мужчины выше 30 % в год. При первой оральной инфекции продукция защитных антител займёт 6 недель, после чего гуморальный иммунитет будет способен защитить организм от повторной генитальной инфекции.
6)Диагностика
Серологический метод. Серологический метод с IgM не позволяет различить антитела к HSV-1 и HSV-2 типам вирусов. Тем не менее, новый иммуноточечный гликопротеин G-специфичный HSV-тест даёт более чем 98 % специфичность и позволяет, таким образом, различить HSV-1 и HSV-2 типы герпеса. Некоторые зарубежные клиницисты считают, что в скором времени тест с IgM будет вытеснен новым.
Цитологический метод. В соскобе поражённого участка эпителия, окрашенного по Романовскому - Гимзе, обнаруживают многоядерные клетки с внутриклеточными включениями.
Вирусологический метод. Заражают культуры клеток и выявляют цитопатологическое действие (ЦПД) в виде гигантских многоядерных клеток с включениями, которые разрушаются. Идентификацию осуществляют в реакции нейтрализации ЦПД, РИФ с моноклональными антителами. На хорионаллантоисной мембране куриных эмбрионов через 2-3 суток образуются белые бляшки.
Биологический метод. При нанесении вирусного материала на скарификацию роговицы кролика возникает кератит, а в мозгу новорождённых мышат - энцефалит.
Иммунофллюоресцентный метод,кожная аллергическая проба, пцр
7)Лечение
На данный момент не существует метода, позволяющего удалить вирус из организма. Антивирусные препараты лишь снижают частоту, продолжительность и тяжесть рецидивов. Анальгетики, например ибупрофен и парацетамол только уменьшают боль и лихорадку. Местные анестетики, такие как прилокаин, лидокаин, бензокаин илитетракаин также снимают зуд и болевые ощущения.
Против герпетических инфекций эффективны: ацикловир, валацикловир, фамцикловир, пенцикловир. Ацикловир был открыт первым, существует множество егодженериков.
Доказано, что ацикловир и валацикловир эффективны при лечении лицевого герпеса (на губах), в том числе у раковых больных. Доказательства в поддержку использования ацикловира при терапии первичного герпетического гингивостоматита, не так сильны.
8)Иммунитет
При первичном инфицировании образуются IgМ-антитела, при рецидивах - IgG и IgА. Вследствие персистенции вируса у инфицированных людей иммунитет является нестерильным и временным - при снижении иммунитета, особенно дефиците естественных киллеров (ЕК), наступает рецидив. Кроме того, вирус герпеса сам способен индуцировать иммунодефицит, одним из механизмов которого является стимуляция синтеза «неэффективных» IgG-антител, которые супрессируют иммунитет, и подавляют ЕК. У людей, резистентных к инфекции, иммунитет осуществляется системой интерферонов, естественных киллеров и Т-киллеров, а также sIgA-антителами. У 80-90 % взрослых имеются IgG-антитела к ВПГ-1. Уровень иммунитета значительно влияет на количество и выраженность рецидивов.
В последние годы в инфекционной патологии все большее значение приобретают герпес-вирусы (от греч. herpes — ползучий). Внимание, которое вирусологи и клиницисты проявляют в последние 25 лет к герпес-вирусным заболеваниям человека, связано с их значительной эпидемиологической ролью и социальной значимостью в современном мире. Неуклонный рост числа герпетических заболеваний у взрослых и детей обусловливает необходимость всестороннего изучения герпетической инфекции и разработки эффективных методов профилактики и лечения разнообразных форм этой инфекции. Среди вирусных инфекций герпес занимает одно из ведущих мест в силу повсеместного распространения вирусов, многообразия клинических проявлений, как правило, хронического течения, а также различных путей передачи вирусов.
Он входит в число наиболее распространенных и плохо контролируемых инфекций человека. Герпес-вирусы могут циркулировать в организме с нормальной иммунной системой бессимптомно, но у людей с иммуносупрессией вызывают тяжелые заболевания со смертельным исходом. По данным ВОЗ, смертность от герпетической инфекции среди вирусных заболеваний находится на втором месте (15,8%) после гепатита (35,8%).
Герпес-вирусы объединены в обширное семейство Herpesviridae и в настоящее время наиболее четко классифицированы. Семейство Herpesviridae включает в себя более 80 представителей, 8 из которых для человека наиболее патогенны (human herpes virus-HHV). Герпес-вирусы — филогенетически древнее семейство крупных ДНК-вирусов — подразделяются на 3 подсемейства в зависимости от типа клеток, в которых протекает инфекционный процесс, характера репродукции вируса, структуры генома, молекулярно-биологических и иммунологических особенностей: α, β и γ ( , по данным Н. Г. Перминова, И. В. Тимофеева и др., Государственный научный центр вирусологии и биотехнологий).
α-герпес-вирусы, включающие HSV-1, HSV-2 и VZV, характеризуются быстрой репликацией вируса и цитопатическим действием на культуры инфицированных клеток. Репродукция α-герпес-вирусов протекает в различных типах клеток, вирусы могут сохраняться в латентной форме, преимущественно в ганглиях.
β-герпес-вирусы видоспецифичны, поражают различные виды клеток, которые при этом увеличиваются в размерах (цитомегалия), могут вызывать иммуносупрессивные состояния. Инфекция может принимать генерализованную или латентную форму, в культуре клеток легко возникает персистентная инфекция. К этой группе относятся CMV, HHV-6, HHV-7.
γ-герпес-вирусы характеризуются тропностью к лимфоидным клеткам (Т- и В-лимфоцитам), в которых они длительно персистируют и которые могут трансформировать, вызывая лимфомы, саркомы. В эту группу входят вирус Эпштейна-Барр и HHV-8-герпес — вирус, ассоциированный с саркомой Капоши (KSHV). KSHV является наиболее близким по геномной организации Т-клеточно-тропному обезьяньему герпес-вирусу Саймири (HVS).
Герпес-вирусы ассоциированы с малигнизацией и способны (по крайней мере, EBV и HVS) трансформировать клетки in vitro. Все герпес-вирусы сходны по морфологическим признакам, размерам, типу нуклеиновой кислоты (двухцепочечная ДНК), икосадельтаэдрическому капсиду, сборка которого происходит в ядре инфицированной клетки, оболочке, типу репродукции, способности вызывать хроническую и латентную инфекцию у человека.
Клонирование герпес-вирусов происходит по следующей схеме: спонтанная случайная адсорбция исходного «материнского» вируса на поверхности клетки-мишени, «раздевание вириона» — расщепление оболочки и капсида, инфильтрация вирусной ДНК в ядро клетки-мишени, формирование и созревание «дочерних» вирионов путем почкования на ядерной мембране. После инфицирования клетки, например вирусом простого герпеса 1 или 2 типов, синтез новых вирусных белков начинается через 2 ч, а число их достигает максимума примерно через 8 ч. В процессе созревания «дочерних» вирионов их оболочки капсиды и ДНК формируются из имеющихся внутри инфицированной клетки аминокислот, белков, липопротеидов и нуклеозидов. Эти молекулы поступают в инфицированную клетку из межтканевых пространств по мере истощения внутриклеточных резервов. В этом отношении вирусы зависят от интенсивности внутриклеточного обмена, который, в свою очередь, определяется природой клетки-мишени. Наиболее высокий темп обмена веществ характерен для короткоживущих клеток эпителиоидного типа, поэтому герпес- вирусы особенно хорошо колонизируют клетки эпителия и слизистых оболочек, крови и лимфатических тканей. Полностью сформированные и готовые к последующей активной репродукции «дочерние» инфекционные вирионы появляются внутри инфицированной клетки через 10 ч, а их число становится максимальным примерно через 15 ч. Количество вирионов в определенной степени влияет на темп распространения инфекции и площадь поражения.
Первая генерация «дочерних» герпес-вирусов начинает поступать в окружающую среду (межклеточные пространства, кровь, лимфу и другие биологические среды) примерно через 18 ч. Это можно пронаблюдать в клинической практике при неконтролируемых процессах (например, при ветряной оспе, опоясывающем герпесе, генерализации цитомегаловирусной инфекции) — элементы герпетической сыпи возникают на коже или слизистых оболочках волнами. В свободном состоянии герпес-вирусы находятся в течение весьма непродолжительного периода (от 1 до 4 ч) — именно такая продолжительность характерна для периода острой интоксикации при герпес-вирусных инфекциях. Срок жизни каждой генерации образовавшихся и адсорбированных герпес-вирусов в среднем составляет 3 сут.
В эпидемиологическом плане наиболее интересны следующие сведения о герпес-вирусах: вирионы чрезвычайно термолабильны — инактивируются при температуре 50-52°С в течение 30 мин, при температуре 37,5°С — в течение 20 ч, устойчивы при температуре 70°С; хорошо переносят лиофилизацию, длительно сохраняются в тканях в 50-процентном растворе глицерина. На металлических поверхностях (монеты, дверные ручки, водопроводные краны) герпес-вирусы выживают в течение 2 ч, на пластике и дереве — до 3 ч, во влажных медицинских вате и марле — до их высыхания при комнатной температуре (до 6 ч).
Уникальными биологическими свойствами всех герпес-вирусов человека является тканевой тропизм, способность к персистенции и латенции в организме инфицированного человека. Персистенция представляет собой способность герпес-вирусов непрерывно или циклично размножаться (реплицироваться) в инфицированных клетках тропных тканей, что создает постоянную угрозу развития инфекционного процесса. Латенция герпес-вирусов — это пожизненное сохранение вирусов в морфологически и иммунохимически видоизмененной форме в нервных клетках регионарных (по отношению к месту внедрения герпес-вируса) ганглиев чувствительных нервов. Штаммы герпес-вирусов обладают неодинаковой способностью к персистенции и латенции и чувствительностью к противогерпетическим препаратам в связи с особенностями их ферментных систем. У каждого герпес-вируса свой темп персистенции и латенции. Среди изучаемых наиболее активны в этом отношении вирусы простого герпеса, наименее — вирус Эпштейна-Барр.
По данным многочисленных исследований, к 18 годам более 90% жителей городов инфицируются одним или несколькими штаммами по меньшей мере 7 клинически значимых герпес-вирусов (простого герпеса 1 и 2 типов, варицелла зостер, цитомегаловирусом, Эпштейна-Барр, герпеса человека 6 и 8 типов). В большинстве случаев первичное и повторное инфицирование происходит воздушно-капельным путем, при прямом контакте или через предметы обихода и гигиены (общие полотенца, носовые платки и т. п.). Доказаны также оральный, генитальный, орогенитальный, трансфузионный, трансплантационный и трансплацентарный пути передачи инфекции.
Герпес-вирусные инфекции широко распространены в мире и имеют тенденцию к неуклонному росту. Особенностью герпес-вирусной инфекции является возможность вовлечения в инфекционный процесс многих органов и систем, чем и обусловлено многообразие вызываемых герпес-вирусами заболеваний, варьирующих от простых кожно-слизистых до угрожающих жизни генерализованных инфекций. Важное свойство герпес-вирусов — это способность после первичного инфицирования в детском возрасте пожизненно персистировать в организме и реактивироваться под влиянием различных экзо- и эндогенных провоцирующих факторов.
Инфицирование человека указанными герпес-вирусами сопровождается клиническими симптомами соответствующего острого инфекционного заболевания в среднем не более чем у 50% людей, в основном у детей: внезапная эритема (вирус герпеса человека 6 типа), афтозный стоматит (вирусы простого герпеса 1 или 2 типов), ветряная оспа (вирус варицелла зостер), инфекционный мононуклеоз (вирус Эпштейна-Барр), мононуклеозоподобный синдром (цитомегаловирус). У остальных пациентов инфекция протекает бессимптомно, что особенно характерно для подростков и взрослых людей. Помимо биологических свойств штамма герпес-вируса, влияние на течение острых и рецидивирующих герпес-вирусных заболеваний оказывают индивидуальные (возрастные, половые, фило- и онкогенетические) особенности иммунного ответа инфицированного человека на многочисленные антигены вируса.
Часто, особенно при снижении иммунореактивности организма, герпес-вирусы выступают в качестве вирусов-оппортунистов, приводя к более тяжелому, с необычными клиническими проявлениями, течению основного заболевания. Вирусы простого герпеса 1 и 2 типов, а также ЦМВ входят в число возбудителей TORCH-инфекций. Они играют важную роль в нарушении репродуктивной функции человека, развитии серьезных заболеваний матери, плода, новорожденного и детей младшего возраста.
Вызываемые вирусами ВПГ, CMV, ВЭБ заболевания рассматриваются как СПИД-индикаторные в связи с их частым обнаружением при данной патологии. В 1988 г. они включены в расширенное определение случаев, подлежащих эпидемиологическому надзору по поводу СПИДа. Результаты исследований последних лет свидетельствуют о роли некоторых герпес-вирусов (ВГЧ-8, ЦМВ, ВЭБ и др.) в развитии ряда злокачественных новообразований: назофарингеальной карциномы, лимфомы Беркитта, В-клеточной лимфомы, рака груди, аденокарциномы кишечника и простаты, карциномы цервикального канала шейки матки, саркомы Капоши, нейробластомы и др.
Наибольшую угрозу для здоровья представляют герпетические нейроинфекции (летальность достигает 20%, а частота инвалидизации — 50%), офтальмогерпес (почти у половины больных приводит к развитию катаракты или глаукомы) и генитальный герпес.
По-видимому, все известные герпес-вирусные инфекции могут рецидивировать, однако порог и причины трансформации острой формы в рецидивирующую для каждого типа герпес-вируса свои. В целом герпес-вирусные инфекции принимают рецидивирующее течение не более чем у 8-20% больных. Рецидивирующие герпес-вирусные заболевания у некоторых людей могут восприниматься как «хронические», когда они развиваются в течение многих лет, не только разрушая физическое здоровье и функции жизненно важных систем, но и психологически крайне неблагоприятно влияя на больного. Поэтому для практических целей герпес-вирусные инфекции классифицируют с учетом одновременно локализации процесса, рецидивирования и этиологии ( ).
Причины рецидивирующего течения герпес-вирусных инфекций многообразны. Одна из них заключается в том, что трансформация острого герпес-вирусного процесса в хронический происходит при явном «попустительстве» иммунной системы. Если приобретенный иммунодефицит в результате химиотерапии или ВИЧ-инфекции легко объясним, то все попытки выяснить, чем обусловлен основной дефект иммунного ответа у иммунологически полноценных людей с рецидивирующим течением герпес-вирусной инфекции, оказались безуспешными. Другая причина заключается, по-видимому, в количественной и качественной особенности персистенции и латенции конкретного штамма герпес-вируса в организме больного.
Диагностика герпетической инфекции
Все методы индикации и идентификации вирусов основаны на следующих принципах:
- выявление вируса per se (электронная микроскопия);
- выявление и идентификация вирусов посредством взаимодействующих с ними клеток (накопление вирусов в чувствительных к ним клетках);
- выявление и идентификация вирусов с помощью антител (МФА, ИФА, РАЛ, ИБ, РН, РСК);
- выявление и идентификация нуклеиновых кислот (ПЦР, МГ).
Электронная микроскопия: быстрая диагностика позволяет обнаружить ГВ или их компоненты непосредственно в пробах, взятых от больного, и дать быстрый ответ через несколько часов. Возбудитель выявляют с помощью электронной микроскопии клинического материала при негативном контрастировании.
Серологические методы уступают по информативности и чувствительности другим способам лабораторной диагностики и не позволяют с достаточной степенью достоверности установить этиологию той или иной формы заболевания. Нарастание титров антител происходит
в поздние сроки (через несколько недель) после заражения или реактивации вируса, и в то же время оно может и не наблюдаться у иммунодефицитных лиц. Для установления 4-кратного нарастания титра антител к герпес-вирусной инфекции (показатель первичной инфекции) необходимо исследование парных сывороток. Серологические реакции (РСК, РН) обладают высокой специфичностью, но относительно низкой чувствительностью, а кроме того, сложны в постановке.
Широкое практическое применение получили иммунофлюоресцентный метод, ИФА, РАЛ, ИБ.
Наиболее точным методом диагностики герпес-вирусной инфекции является выделение вируса из различных клеточных культур.
Для обнаружения вируса герпеса используют молекулярно-биологические методы: полимеразную цепную реакцию и реакцию молекулярной гибридизации, которые позволяют выявить наличие вирусной нуклеиновой кислоты в исследуемом материале. ПЦР может считаться самой чувствительной и быстрой реакцией. Чувствительность метода дает возможность определить одну молекулу искомой ДНК в образцах, содержащих 10 клеток.
Лечение герпетической инфекции
Лечение герпетической инфекции до настоящего времени остается сложной задачей. Хроническое течение процесса приводит к иммунной перестройке организма: развитию вторичной иммунной недостаточности, угнетению реакции клеточного иммунитета, снижению неспецифической защиты организма. Несмотря на разнообразие лекарственных препаратов, использующихся для лечения герпетической инфекции, лекарственных средств, обеспечивающих полное излечение от герпеса, не существует. Герпес-вирусная инфекция относится к трудно контролируемым заболеваниям. Это связано, в первую очередь, с разнообразием клинических поражений, развитием резистентности вируса к лекарственным средствам, наличием у герпес-вирусов молекулярной мимикрии. Поэтому для успешного лечения герпетической инфекции необходимо правильно подобрать противовирусный препарат, его дозу и длительность лечения, использовать комбинацию различных лекарств. В схемы терапии для повышения эффективности лечения необходимо также включать иммунобиологические препараты, способствующие коррекции иммунного статуса, а также патогенетические средства, облегчающие состояние пациента.
В настоящее время все антигерпетические средства подразделяются на 3 основные группы антивирусных препаратов ( ).
Механизм действия химиопрепаратов (аномальных нуклеозидов: валтрекса, вектавира, фамвира, цимевена) связан с угнетением синтеза вирусной ДНК и репликации вирусов путем конкурентного ингибирования вирусной ДНК-полимеразы.
В препаратах-иммуномодуляторах (алпизарин, имунофан, ликопид, полиоксидоний) активно действующие вещества обладают иммуностимулирующими свойствами в отношении клеточного и гуморального иммунитетов, окислительно-восстановительных процессов, синтеза цитокинов.
Препараты-индукторы ИФН (амиксин, неовир, циклоферон) сочетают этиотропный и иммуномодулирующий эффекты действия. Препараты индуцируют образование эндогенных ИФН (α, β, γ) Т- и В-лимфоцитами, энтероцитами, гепатоцитами.
Особое место среди средств антигерпес-вирусной терапии занимает герпетическая вакцина для активации клеточного иммунитета, его иммунокоррекции в фазе ремиссии. Вакцинация преследует 2 цели: предупреждение первичной инфекции и возникновения состояния латентности, а также предупреждение или облегчение течения заболевания.
Однако, несмотря на наличие обширного списка антигерпетических препаратов, герпес остается до сих пор плохо контролируемой инфекцией. Это обусловлено и генотипическими особенностями возбудителя, и длительной персистенцией вируса в организме, и формированием штаммов, устойчивых к антивирусным препаратам. Максимальный клинический эффект может быть получен только при рациональной комплексной терапии лекарственных средств с различным механизмом действия.
Санкт-Петербургской группой ученых вирусологов и инфекционистов во главе с В. А. Исаковым предложена программа лечения и профилактики герпетической инфекции (табл. 4).
Преимущества комплексной терапии ГИ.
- Сочетанное применение противогерпетических химиопрепаратов и иммунобиологических средств обеспечивает синергидный эффект.
- Благодаря снижению дозы противовирусного ХПП уменьшается вероятность развития побочных эффектов, сокращается токсическое воздействие на организм больного.
- Снижается вероятность возникновения устойчивых штаммов герпес-вирусов к данному препарату.
- Достигается иммунокорригирующий эффект.
- Сокращается продолжительность острого периода болезни и сроков лечения.
Таким образом, терапия ГИ является сложной и многокомпонентной задачей.
По вопросам литературы обращайтесь в редакцию.
Т. К. Кускова,
кандидат медицинских наук
Е. Г. Белова
, кандидат медицинских наук
МГМСУ, Москва
1. Вирусология. История вирусологии. Шамберлан. Ру. Пастер. Ивановский.
2. Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
3. Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
4. Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
5. Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
7. Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
8. Генетика вирусов. Характеристика вирусных популяций. Генофонд вирусных популяций.
9. Мутации вирусов. Спонтанные мутации вирусов. Индуцированные мутации вирусов. Проявление мутаций вирусов в фенотипе.
10. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами. Обмен фрагментами генома вирусами. Антигенный шифт.
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко.
Ранняя стадия репродукции герпесвирусов . В ранней стадии синтезируются «ранние белки», кодируемые проксимальной третью молекулы ДНК. Они проявляют регуляторные свойства, включая активацию транскрипции других участков вирусного генома, кодирующих ДНК-полимеразу и ДНК-связывающие белки.
Поздняя стадия репродукции герпесвирусов . В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов).
Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов - формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
Поксвирусы. Репродукция поксвирусов.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью осуществляется вирусными полимеразами.
Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз, так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНК-полимеразу, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий.
В репродуктивном цикле выделяют три стадии- начальную, раннюю и позднюю.
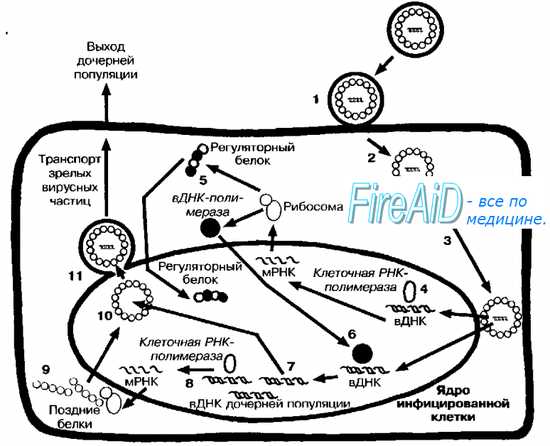 Рис. 5-4. Репликативный цикл ДНК-содержащих вирусов
(на примере репродукции вируса герпеса). После адсорбции (1) вирус проникает в клетку путём слияния с мембраной (2). Нуклеокапсид транспортируется к ядерной оболочке (3), и вирусная ДНК (вДНК) проникает в ядро клетки, где начинается её транскрипция клеточной ДНК-зависимой РНК-полимеразой (4). Вначале транскрибируются «ранние гены». В результате трансляции «ранней порции» вирусного генома синтезируются «ранние белки» (5), включая регуляторные, матричные и вирусные полимеразы. Вирусная полимераза проникает в геном клетки (6), где запускает синтез молекул ДНК дочерних популяций (7). Часть вирусной ДНК («поздние гены») дочерних популяций транскрибируется клеточной РНК-полимеразой (8), что приводит к синтезу «поздних белков» (9), необходимых для сборки дочерних популяций (10). Последние покидают ядро, отпочковываясь от его мембраны (11), фрагменты которой содержат в составе своей обопочки.
Рис. 5-4. Репликативный цикл ДНК-содержащих вирусов
(на примере репродукции вируса герпеса). После адсорбции (1) вирус проникает в клетку путём слияния с мембраной (2). Нуклеокапсид транспортируется к ядерной оболочке (3), и вирусная ДНК (вДНК) проникает в ядро клетки, где начинается её транскрипция клеточной ДНК-зависимой РНК-полимеразой (4). Вначале транскрибируются «ранние гены». В результате трансляции «ранней порции» вирусного генома синтезируются «ранние белки» (5), включая регуляторные, матричные и вирусные полимеразы. Вирусная полимераза проникает в геном клетки (6), где запускает синтез молекул ДНК дочерних популяций (7). Часть вирусной ДНК («поздние гены») дочерних популяций транскрибируется клеточной РНК-полимеразой (8), что приводит к синтезу «поздних белков» (9), необходимых для сборки дочерних популяций (10). Последние покидают ядро, отпочковываясь от его мембраны (11), фрагменты которой содержат в составе своей обопочки.
Начальная стадия репродукции поксвирусов запускается сразу же после раздевания вируса и выхода вирусной ДНК в цитоплазму.
Ранняя стадия репродукции поксвирусов . На этом этапе транскрибируется примерно половина вирусной ДНК. Синтезируются ферменты, кодируемые «ранними генами» и участвующие в репликации вирусной ДНК. Параллельно в небольшом количестве образуются структурные белки.
Поздняя стадия репродукции поксвирусов совпадает с началом репликации ДНК (что переключает механизмы транскрипции на считывание второй половины генома). Регуляторные белки блокируют трансляцию «ранней мРНК» и запускают синтез поздних (структурных) белков. Сборка вирионов осуществляется только в цитоплазме посредством реакций мембранного синтеза. Высвобождение зрелых популяций сопровождается лизисом клетки.