زیست توده، تولید اولیه فیتوپلانکتون و کلروفیل. عوامل موثر بر توسعه فیتوپلانکتون لیست پیشنهادی پایان نامه ها
V.A. چوگاینووا، آی.یو. Makedonskaya شعبه شمالی PINRO، آرخانگلسک، روسیه ایمیل: [ایمیل محافظت شده]
تولید اولیه، که بیشترین سهم را جلبکهای پلانکتون در آن میکنند، همراه با مواد آلی آلوکتونی که وارد مخزن میشوند، پایه مادی و انرژی تمام مراحل بعدی فرآیند تولید را تشکیل میدهند.
به لطف توسعه روش هایی برای مطالعه تولید اولیه، بهره وری بیولوژیکی کلی یک مخزن بیان کمی دریافت کرده است. نیاز به توصیف کمی مواد آلی سنتز شده در طول فتوسنتز پلانکتون به وضوح هنگام حل بسیاری از مسائل و شیوه های هیدروبیولوژی ظاهر می شود. اما، علیرغم این، دانش ویژگیهای تولید فیتوپلانکتونها چیزهای زیادی برای دلخواه باقی میگذارد.
مواد و روش ها
مطالعات مربوط به بهره وری اولیه فیتوپلانکتون ها از 7 ژوئیه تا 21 ژوئیه 2007 در ایستگاه SevPINRO در تنگه Pechakovskaya Salma (جزایر Solovetsky) در ایستگاه های روزانه (در مجموع 14 تعیین بهره وری ناخالص اولیه) انجام شد. همچنین یکی از اهداف تحقیق ما تغییرات کمی و کیفی روزانه در فیتوپلانکتون بود. در این راستا 13 نمونه فیتوپلانکتون در لایه سطحی منطقه ساحلی جمع آوری و فرآوری شد. نمونه های فیتوپلانکتون در تاریخ 14 تا 15 تیرماه پس از دو ساعت برداشته شد. مجموعه مشاهدات، همراه با شاخص های کمی و کیفی فیتوپلانکتون، شامل تعیین دما، شوری آب و میزان اکسیژن بود.
تجزیه و تحلیل نمونههای آب دریا با استفاده از روشهای پذیرفتهشده در عمل هیدروشیمی انجام شد. اکسیژن محلول در آب با روش حجمی وینکلر (Manual..., 2003) تعیین شد. انکوباسیون نمونه های آب برای تعیین شدت فتوسنتز در فلاسک های تیره و روشن در دمای آب دریا و نور طبیعی انجام شد. نمونه های ترکیب رنگدانه فیتوپلانکتون از طریق فیلترهای غشایی Vladipor با قطر 35 میلی متر و اندازه منافذ 0.65 میکرون فیلتر شدند. نمونه های فیلتر در فریزر در ظرف حاوی سیلیکاژل نگهداری شدند. رنگدانه های ریزجلبک در شرایط آزمایشگاهی با استفاده از روش های استاندارد تعیین شدند. چگالی نوری عصاره در طول موج های 480، 630، 647، 664 و 750 نانومتر اندازه گیری شد. غلظت کلروفیل "a" با استفاده از فرمول های جفری و هامفری محاسبه شد (Jeffrey S.W., Humphrey G.F., 1975).
در نتیجه تحقیق، تعدادی مشاهدات در مورد تغییرات تولید ناخالص اولیه در طی 15 روز به دست آمد و پویایی روزانه شاخص های کمی و کیفی فیتوپلانکتون شناسایی شد.
نتایج و بحث آن
با توجه به مشاهدات ما، مقادیر تولید ناخالص اولیه (PPtotal) در Pechakovskaya Salma در طیف گسترده ای متفاوت است - 0.33-1.65 mgO2 / L / روز (که مربوط به 124-619 mgC / m3 / روز است)، مقدار متوسط. mgO2/l/day 63/0 (mgC/m3/day 4/256) بود. حداکثر مقادیر در دو روز گذشته ثبت شد که احتمالاً به دلیل شرایط آب و هوایی مطلوب تر است (شکل 1). این مقادیر اساساً با PPtotal به دست آمده در سال های گذشته در این زمینه مطابقت دارد (چوگاینووا، Makedonskaya، 2007).
به طور کلی، فتوسنتز ناخالص در طی 15 روز کاملاً یکنواخت تغییر کرد و یک ویژگی موج مانند را نشان داد.
لازم به ذکر است که شرایط طبیعی پایدار در این مدت مشاهده شد. بنابراین، دمای آب در محدوده 8.4 درجه سانتیگراد در ابتدای مشاهدات تا 10 درجه سانتیگراد در پایان (میانگین 9.66 درجه سانتیگراد)، شوری در محدوده 26.2-26.9٪٪ (با میانگین 26.6٪) متغیر بود. o). شرایط آب و هوایی نیز تقریبا یکنواخت بود.
شاخص های تخریب در کل دوره مشاهده از PPTotal فراتر رفت و فقط در پایان دوره مقادیر آنها نزدیک به تعادل بود. به طور متوسط، تخریب 414.4 mg C / m 3 / روز (با تغییر 86.3 - 742.5 mg C / m 3 / روز) بود.
تغییرات روزانه در فراوانی، زیست توده، و مجموعه گونه های فیتوپلانکتون غالب در معرض برخی از نوسانات بود. زیست توده از 94.8 تا 496.44 میکروگرم در لیتر، و فراوانی - از 4860 تا 18220 سلول در لیتر (شکل 2) متغیر بود.میانگین روزانه فراوانی و زیست توده به ترتیب 10277 سلول در لیتر و 311.21 میکروگرم در لیتر بود.

تعداد کل گونه های ریزجلبک در نمونه های جولای از 13 تا 25 در طول روز در نوسان بود که در مجموع 45 گونه در طول مطالعه کشف شد. مجموعه گونه های غالب فیتوپلانکتون های جولای شامل: کریپتوفیت ها - Leoucocryptos marina. سبز - Piramimonas sp.، Chlorococales کوچک؛ دیاتومها - Thalassiosira Nordenscioldii، Leptocylindrus danicus، Detonula confervacea، L icmophora paradoxa. دینوفیت ها - Gymnodinium arcticum. مجموعه ریزجلبکها در فصل تابستان برای این منطقه بسیار رایج است (Makedonskaya، 2007).
نقش اصلی در فرآیند فتوسنتز توسط کلروفیل "a" ایفا می شود؛ همه رنگدانه های دیگر فقط انرژی جذب شده را به کلروفیل "a" منتقل می کنند. بنابراین، محتوای کلروفیل "a" مهمترین مشخصه فعالیت فتوسنتزی فیتوپلانکتون است، که از آن می توان شاخص های شماتیک زیست توده فیتوپلانکتون را نیز تعیین کرد (شکل 2 را ببینید). مطالعه روابط کمی بین رنگدانه های مختلف فیتوپلانکتون به ما اجازه می دهد تا در مورد غلبه گروه خاصی از جلبک ها در آب دریا قضاوت کنیم. بنابراین، بخش عمده ای از فیتوپلانکتون های دریایی از دیاتوم ها و جلبک های پریدینیم تشکیل شده است که حاوی کلروفیل های "a" و "c" هستند. تعریف حتی کوچک
مقدار کلروفیل "b" نشان دهنده رشد جلبک های کوچک تاژک دار (سبز) و آبی-سبز است. روابط بین رنگدانه های جلبک نیز وضعیت فیزیولوژیکی جمعیت فیتوپلانکتون را مشخص می کند. کلروفیل "a" 51 درصد از فیتوپیگمنت ها را تشکیل می دهد. کلروفیل «b» موجود در کلروپلاست جلبکهای سبز 24 درصد است، سهم کلروفیل «c» که در سلولهای دیاتومها، دینوفیتها و سایر بخشهای جلبک یافت میشود، 25 درصد است. این نسبت رنگدانه ها نشان دهنده فعالیت فتوسنتزی شدید فیتوپلانکتون ها است. این به طور غیرمستقیم با اشباع اکسیژن آب ها، که در طول ایستگاه روزانه 110-130٪ نشسته بود، و همچنین توسط شاخص های PPtotal تأیید می شود.
تلاش برای مقایسه مقادیر کلروفیل ها، زیست توده و فراوانی ریزجلبک ها با چرخه جزر و مدی نشان داد که غلظت آنها به فاز جزر و مد بستگی ندارد. و با محتوای و اشباع آب از اکسیژن در آنتی فاز هستند.
در تابستان، در منطقه تنگه Pechakovskaya Salma، مقادیر بالایی از تولید فیتوپلانکتون اولیه، قابل مقایسه با بهار، مشاهده شد.
تغییرات در ترکیب کیفی و کمی فیتوپلانکتون ها در طول روز تفاوت آشکاری ندارد. دلیل این امر، به احتمال زیاد، رژیم هیدرولوژیکی و هیدروشیمیایی نسبتاً پایدار آبهای Pechakovskaya Salma در طول دوره مشاهده است.
تحقیقات بیشتری برای روشن شدن تغییرات روزانه و فصلی در جامعه فیتوپلانکتون در این منطقه مورد نیاز خواهد بود.
ادبیات
Makedonskaya I.Yu.، 2007. در مورد پویایی فصلی و بین سالانه فیتوپلانکتون در Pechakovskaya Salma خلیج Onega دریای سفید // مشکلات مطالعه، استفاده منطقی و حفاظت از منابع طبیعی دریای سفید - مواد X کنفرانس بین المللی. آرخانگلسک. ص 154-158.
Chugainova V.A., Makedonskaya I.Yu., 2008. تغییرات فصلی در بهره وری اولیه و کلروفیل "a" در منطقه جزایر سولووتسکی. //اکوسیستم های ساحلی دریایی: جلبک ها، بی مهرگان و محصولات آنها. چکیده گزارش های سومین کنفرانس بین المللی علمی و کاربردی. ولادی وستوک: مرکز TINRO. ص 163-164.
رهنمودهایی برای تجزیه و تحلیل شیمیایی دریا و آب های شیرین در هنگام پایش زیست محیطی مخازن ماهیگیری و مناطق ماهیگیری امیدوار کننده اقیانوس جهانی.، 2003. M.: انتشارات VNIRO. -202 s.
Jeffrey S.W., Humphrey G.F., 1975. معادلات اسپکتروفتومتری جدید برای تعیین کلروفیل a، b، c1 و c2 در گیاهان عالی، جلبک ها و فیتوپلانکتون های طبیعی // بیوشیمی. و فیزیول. پفلانز. Bd. 167. شماره 2. ص 191-194.
کارایی اولیه فیتوپلانکتون در کمربند PECHAKOVSKAJA SALMA دریای سفید در طول دوره تابستان
V.A. چوگاینووا، آی.جی. Makedonskaya
شعبه شمالی PINRO، آرخانگلسک، روسیه ایمیل: [ایمیل محافظت شده]
بررسی کارایی اولیه فیتوپلانکتون در 7-21 ژوئیه 2007 در استقرار دائمی SevPINRO در کمربند Pechakovskaja Salma، (جزایر Solovetskiye) در ایستگاههای روزانه (14 تعریف از بازده اولیه کل در کل) انجام شد. بر اساس مشاهدات ما، مقادیر کل تولید اولیه
(TPP) در Pechakovskaja Salma در طیف وسیعی متفاوت است - 0.33-1.65 mgO 2 / L / روز (که مربوط به 124 - 619 mg C / m 3 / روز است)، مقدار متوسط 0.63 mgO 2 / L / روز (256.4 mg C / روز) است. متر 3 / روز). به طور کلی، فتوسنتز کل در عرض 15 روز در فواصل منظم به اندازه کافی تغییر کرد و ویژگی موجی را نشان داد. تغییرات روزانه در تعداد، زیست توده و مجموعه ای از گونه های فیتوپلانکتون غالب نیز در معرض برخی نوسانات قرار گرفتند. زیست توده در محدوده های 94.8 تا 496.44 میلی گرم بر لیتر و تعداد - از 4860 تا 18220 سلول در لیتر تغییر کرد. میانگین روزانه تعداد و زیست توده به ترتیب 10277 سلول در لیتر و 311.21 میلیگرم در لیتر است.
فیتوپلانکتون، با اتصال CO 2 در طول فتوسنتز و تشکیل مواد آلی، تمام زنجیره های غذایی در اقیانوس را به وجود می آورد. تجزیه و تحلیل انواع داده ها در مورد میزان فیتوپلانکتون در مناطق مختلف اقیانوس جهانی (از پایان قرن نوزدهم، محاسبه شده از برآوردهای شفافیت موجود، و از اوایل دهه 1980، به دست آمده از راه دور از فضاپیماها) نشان می دهد که زیست توده آن کاهش یافته است. در طول قرن گذشته با نرخی در حدود 1% در سال. قابل توجه ترین کاهش برای مناطق الیگوتروف مرکزی اقیانوس مشاهده شد. اگرچه این مناطق با بهره وری بسیار پایین مشخص می شوند، اما منطقه عظیمی را اشغال می کنند و بنابراین سهم کلی آنها در تولید و زیست توده فیتوپلانکتون اقیانوس ها بسیار قابل توجه است. محتمل ترین دلیل کاهش زیست توده افزایش دمای لایه سطحی اقیانوس است که منجر به کاهش عمق اختلاط و کاهش عرضه عناصر غذایی معدنی از لایه های زیرین می شود.
حدود نیمی از کل تولید اولیه سیاره ما (یعنی مواد آلی تولید شده توسط گیاهان سبز و سایر موجودات فتوسنتزی کننده) از اقیانوس حاصل می شود. تولیدکنندگان اصلی اقیانوس جلبکهای میکروسکوپی و سیانوباکتریهای معلق در لایههای بالایی ستون آب (که در مجموع فیتوپلانکتون نامیده میشود) هستند. مطالعات کمی در مقیاس بزرگ در مورد تولید و زیست توده فیتوپلانکتون در اقیانوس جهانی در دهه 1960 و 70 آغاز شد. سپس محققان (از جمله کسانی از مؤسسه اقیانوسشناسی آکادمی علوم اتحاد جماهیر شوروی) به روشی مبتنی بر جذب ایزوتوپ کربن رادیواکتیو 14 C توسط فیتوپلانکتونها تکیه کردند. روی کشتی بلند شد. در نتیجه این کارها، نقشه هایی از توزیع فیتوپلانکتون در سراسر اقیانوس جهانی ساخته شد (به عنوان مثال نگاه کنید به: Koblentz-Mishke et al., 1970). در نواحی مرکزی و وسیع اقیانوس، زیست توده فیتوپلانکتون و تولید آن بسیار کم است. مقادیر بالای زیست توده و تولید محدود به مناطق ساحلی و بالارونده است (نگاه کنید به: بالا آمدن)، جایی که آب های عمیق غنی از عناصر غذایی معدنی به سطح می آیند. اول از همه، اینها فسفر و نیتروژن هستند که کمبود آنها رشد فیتوپلانکتون ها را در بیشتر آب های اقیانوسی محدود می کند.
مرحله جدیدی در مطالعه کمی توزیع فیتوپلانکتون در اقیانوس جهانی در اواخر دهه 1970 و پس از ظهور روش های دور (از ماهواره ها) برای سنجش آب های سطحی و تعیین محتوای کلروفیل در آنها آغاز شد. اگرچه بیش از 10 درصد از فوتون های نور که از آب منعکس می شود و حاوی اطلاعاتی در مورد رنگ آن است، به دستگاه های واقع در مرز بالایی جو نمی رسد، اما برای محاسبه مقدار کلروفیل کافی است و بر این اساس، زیست توده فیتوپلانکتون (شکل 1). مقادیر زیست توده همچنین می تواند برای قضاوت در مورد تولید فیتوپلانکتون مورد استفاده قرار گیرد، که در طول مطالعات ویژه مقایسه داده های ماهواره ای با نتایج تخمین های تولید به دست آمده به صورت تجربی تأیید شد. در موقعیتروی کشتی های تحقیقاتی البته، دستگاه های مختلف داده های کمی متفاوت ارائه می دهند، اما تصویر کلی از توزیع فضایی فیتوپلانکتون و پویایی آن (فصلی و بین سالانه) بسیار دقیق است. کافی است که بگوییم دستگاه Sea WiFS (سنسور میدان دید گسترده دریا) در عرض دو روز تمام اقیانوس های جهان را اسکن می کند.
حجم عظیمی از داده های انباشته شده در 30 سال گذشته، شناسایی نوسانات دوره ای خاصی را در زیست توده فیتوپلانکتون، به ویژه مرتبط با ال نینو، یا به طور دقیق تر، با "نوسانات جنوبی" (El Niño-Southern Oscillation) ممکن کرده است. با تجزیه و تحلیل این مواد، محققان وجود تغییرات طولانیمدت در زیست توده فیتوپلانکتونها را پیشنهاد کردند، اما به دلیل کمبود دادهها برای دوره قبل از اندازهگیریهای ماهوارهای، شناسایی آنها دشوار بود. اخیراً متخصصان دانشگاه دالهوسی کانادا در هالیفاکس (دانشگاه دالهوسی، هالیفاکس، نوا اسکوشیا) تلاشی برای حداقل تا حدی حل این مشکل انجام دادند. زیست توده فیتوپلانکتون 50 و حتی 100 سال پیش را می توان با تخمین های شفافیت ارزیابی کرد، مقداری که به طور منظم در اکتشافات تحقیقاتی از پایان قرن 19 اندازه گیری می شود.

ابزاری برای اندازه گیری شفافیت آب، بسیار ساده اما بسیار مفید، در سال 1865 توسط ستاره شناس ایتالیایی (و در عین حال کشیش) آنجلو سکی، که وظیفه ترسیم نقشه شفافیت دریای مدیترانه را بر عهده داشت، اختراع شد. ناوگان پاپ دستگاهی که «دیسک سچی» نامیده می شود (شکل 2 را ببینید)، یک صفحه فلزی سفید رنگ به قطر 20 یا 30 سانتی متر است که روی یک طناب مشخص شده در آب فرو می رود. عمقی که ناظر دیگر دیسک را نمی بیند، شفافیت Secchi است. از آنجایی که بخش اصلی ماده معلق که بر شفافیت آب تأثیر می گذارد، فیتوپلانکتون است، هر گونه تغییر در مقدار شفافیت. به عنوان یک قاعده، آنها به خوبی تغییرات در مقدار فیتوپلانکتون را منعکس می کنند.

بر اساس برآوردهای استاندارد شده شفافیت موجود از سال 1899، و بر اساس نتایج مقایسه اخیر مقادیر شفافیت با غلظت کلروفیل، محققان ابتدا تصویری از توزیع زیست توده فیتوپلانکتون در اقیانوس ها به دست آوردند (شکل 3) و دوم، تغییرات در زیست توده فیتوپلانکتون در طول یک دوره صد ساله (شکل 4). در مجموع، آنها نتایج بیش از 455 هزار اندازه گیری را در اختیار داشتند که دوره 1899 تا 2008 را پوشش می دهد. در عین حال، داده های مربوط به منطقه ساحلی (کمتر از 1 کیلومتر از ساحل و در اعماق کمتر از 25 متر) عمداً در نمونه گنجانده نشد، زیرا در چنین مکان هایی تأثیر رواناب از ساحل بسیار قابل توجه است. . بیشتر اندازه گیری ها پس از سال 1930 در مناطق شمالی اقیانوس اطلس و اقیانوس آرام انجام شد. نتیجه اصلی که نویسندگان به آن می رسند کاهش تدریجی زیست توده کل فیتوپلانکتون در طول قرن گذشته با نرخ متوسط حدود 1٪ در سال است.

برای ارزیابی روندهای محلی، کل منطقه آبی اقیانوس جهانی به شبکه ای با سلول هایی با ابعاد 10 × 10 درجه تقسیم شد و همه مقادیر به عنوان میانگین در هر سلول محاسبه شد. کاهش در زیست توده فیتوپلانکتون در 59٪ از سلول هایی که داده های به اندازه کافی قابل اعتماد در دسترس بود مشاهده شد. بیشتر این سلول ها در عرض های جغرافیایی بالا (بیش از 60 درجه عرض جغرافیایی) قرار دارند. با این حال، برای برخی از مناطق اقیانوس، افزایش زیست توده مشاهده شد - به ویژه، در بخش شرقی اقیانوس آرام، و همچنین در مناطق شمالی و جنوبی اقیانوس هند. مناطق اولیگوتروف مرکزی اقیانوس ها در واقع مناطق آبی اشغالی را گسترش داده اند و در این مناطق با وجود بهره وری پایین، هم اکنون حدود 75 درصد از کل تولید اولیه اقیانوس جهانی تشکیل می شود.
برای تصور تغییرات در سطح مناطق بزرگ، کل منطقه اقیانوس به 10 منطقه تقسیم شد (شکل 5): قطب شمال، شمال، استوایی و اقیانوس اطلس جنوبی، بخش های شمالی و جنوبی اقیانوس هند، شمال، استوایی و اقیانوس آرام جنوبی و اقیانوس جنوبی. تجزیه و تحلیل داده های میانگین برای این مناطق بزرگ نشان داد که افزایش قابل توجهی فقط برای بخش جنوبی اقیانوس هند و افزایش آماری غیر قابل اعتماد برای بخش شمالی اقیانوس هند مشاهده شد. برای تمام مناطق دیگر، کاهش قابل توجهی در زیست توده فیتوپلانکتون مشاهده شد.

با بحث در مورد دلایل احتمالی تغییرات مشاهده شده، نویسندگان در درجه اول به افزایش دمای لایه سطحی ستون آب توجه می کنند. تقریباً تمام اقیانوس را پوشانده و منجر به کاهش ضخامت لایه مخلوط شده است. بر این اساس، هجوم عناصر غذایی معدنی (عمدتاً فسفات ها و نیترات ها) از لایه های زیرین کاهش می یابد. با این حال، نویسندگان اذعان می کنند که چنین توضیحی برای عرض های جغرافیایی بالا مناسب نیست. در آنجا، گرم شدن لایه بالایی باید به جای کاهش، تولید فیتوپلانکتون و زیست توده را افزایش دهد. واضح است که مکانیسم هایی که تغییرات در مقیاس بزرگ در زیست توده فیتوپلانکتون را تعیین می کنند، نیاز به مطالعه بیشتر دارند.
گیاهان مخازن ما: فیتوپلانکتون
فیتوپلانکتون ها جلبک های میکروسکوپی هستند که آزادانه در ستون آب شناور هستند. برای زندگی در این حالت، در طول فرآیند تکامل، آنها تعدادی سازگاری ایجاد کرده اند که به کاهش تراکم نسبی سلول ها (انباشته شدن آخال ها، تشکیل حباب های گاز) و افزایش اصطکاک آنها (فرآیندهای اشکال مختلف، رشد) کمک می کند.
فیتوپلانکتون آب شیرین عمدتاً توسط جلبک های سبز، آبی-سبز، دیاتومه ها، پیروفیت ها، جلبک های طلایی و اوگلنا نشان داده می شود.
توسعه جوامع فیتوپلانکتون با دوره ای مشخص رخ می دهد و به عوامل مختلفی بستگی دارد. به عنوان مثال، افزایش زیست توده ریز جلبک ها تا یک نقطه خاص به نسبت مقدار نور جذب شده اتفاق می افتد. جلبکهای سبز و آبی-سبز تحت نور 24 ساعته بیشتر تولید مثل میکنند، در حالی که دیاتومها در دورههای نوری کوتاهتر تولید مثل میکنند. شروع فصل رشد فیتوپلانکتون در ماه مارس تا آوریل تا حد زیادی با افزایش دمای آب همراه است. دیاتومه ها دمای بهینه پایینی دارند، در حالی که سبزها و سبز آبی ها دارای بهینه بالاتری هستند. بنابراین، در بهار و پاییز، زمانی که دمای آب بین 4 تا 15 درجه است، دیاتوم ها در بدنه های آبی غالب هستند. افزایش کدورت آب ناشی از سوسپانسیونهای معدنی، شدت رشد فیتوپلانکتونها را کاهش میدهد، به ویژه آنهایی که سبز-آبی هستند. دیاتوم ها و جلبک های پروتوکوکی نسبت به افزایش کدورت آب حساسیت کمتری دارند. در آب های غنی از نیترات، فسفات و سیلیکات ها، عمدتاً دیاتوم ها رشد می کنند، در حالی که سبز و سبز آبی از نظر محتوای این مواد مغذی تقاضای کمتری دارند.
ترکیب گونهای و فراوانی فیتوپلانکتونها نیز تحتتاثیر محصولات متابولیکی خود جلبکها قرار میگیرد، بنابراین، همانطور که در ادبیات علمی اشاره شد، روابط متضاد بین برخی از آنها وجود دارد.
از بین گونههای مختلف فیتوپلانکتونهای آب شیرین، دیاتومها، جلبکهای سبز و آبی-سبز بیشترین و به ویژه برای غذا ارزشمند هستند.
سلول های دیاتومه ها مجهز به یک پوسته دوگوشی ساخته شده از سیلیس هستند. خوشه های آنها با رنگ مشخص قهوه ای مایل به زرد متمایز می شوند. این میکروفیت ها نقش مهمی در تغذیه زئوپلانکتون ها دارند، اما به دلیل محتوای کم مواد آلی ارزش غذایی آنها به اندازه جلبک های پروتوکوکی قابل توجه نیست.
ویژگی بارز جلبک سبز رنگ سبز معمولی آن است. سلول های آنها که حاوی یک هسته و یک کروماتوفور است، شکل متفاوتی دارند و اغلب مجهز به خار و مو هستند. برخی چشم قرمز دارند (انگ). در میان نمایندگان این بخش، جلبک های پروتوکوکی اشیاء کشت انبوه هستند (کلرلا، سندسموس، آنکیسترودسموس). سلول های آنها از نظر اندازه میکروسکوپی هستند و به راحتی برای موجودات آبزی تغذیه کننده از فیلتر قابل دسترسی هستند. محتوای کالری ماده خشک این جلبک ها نزدیک به ۷ کیلو کالری در گرم است. آنها حاوی مقدار زیادی چربی، کربوهیدرات و ویتامین هستند.
سلول های جلبک سبز آبی کروماتوفور یا هسته ندارند و به طور یکنواخت به رنگ سبز آبی هستند. گاهی اوقات رنگ آنها می تواند بنفش، صورتی و سایه های دیگر به خود بگیرد. محتوای کالری ماده خشک به 5.4 کیلو کالری در گرم می رسد. این پروتئین از نظر ترکیب اسید آمینه کامل است، اما به دلیل حلالیت کم آن برای ماهی غیرقابل دسترسی است.
فیتوپلانکتون نقش کلیدی در ایجاد منبع غذایی طبیعی مخازن دارد. میکروفیت ها به عنوان تولیدکنندگان اولیه، با جذب ترکیبات معدنی، مواد آلی را سنتز می کنند که توسط زئوپلانکتون (مصرف کننده اولیه) و ماهی (مصرف کننده ثانویه) استفاده می شود. ساختار زئوپلانکتون تا حد زیادی به نسبت اشکال بزرگ و کوچک در فیتوپلانکتون بستگی دارد.
یکی از عوامل محدود کننده رشد میکروفیت ها محتوای نیتروژن محلول (عمدتا آمونیوم) و فسفر در آب است. برای استخرها، هنجار بهینه 2 میلی گرم N/L و 0.5 میلی گرم فسفر در لیتر است. افزایش زیست توده فیتوپلانکتون با کاربرد جزئی 1 سی در هکتار نیتروژن فسفر و کودهای آلی در هر فصل تسهیل می شود.
پتانسیل تولید جلبک بسیار زیاد است. با استفاده از فناوری مناسب می توان تا 100 تن ماده خشک کلرلا را از 1 هکتار سطح آب به دست آورد.
کشت صنعتی جلبک شامل تعدادی مراحل متوالی با استفاده از انواع مختلف راکتورها (کلتیواتور) در محیط های مایع می باشد. میانگین بازده جلبک طبق VNIIPRKh از 2 تا 18.5 گرم ماده خشک در هر متر مربع در روز متغیر است.
اندازه گیری بهره وری فیتوپلانکتون، میزان تشکیل مواد آلی در طول فتوسنتز است. جلبک ها منبع اصلی تولید اولیه هستند. تولید اولیه مقدار ماده آلی سنتز شده توسط موجودات اوتروفیک در واحد زمان است که معمولاً بر حسب کیلوکالری بر متر در روز بیان می شود.
فیتوپلاکتون با بیشترین دقت، سطح تغذیه ای یک مخزن را تعیین می کند. به عنوان مثال، آب های اولیگوتروف و مزوتروف با نسبت کم فراوانی فیتوپلانکتون به زیست توده آن مشخص می شوند، در حالی که آب های هیپرتروفیک با نسبت بالایی مشخص می شوند. زیست توده فیتوپلانکتون در مخازن هیپرتروفیک بیش از 400 میلی گرم در لیتر، در اوتروفیک - 40.1-400 میلی گرم در لیتر، در دیستروفیک - 0.5-1 میلی گرم در لیتر است.
اوتروفیکاسیون انسانی - افزایش اشباع یک مخزن با مواد مغذی - یکی از مشکلات اساسی است. درجه فعالیت فرآیندهای بیولوژیکی در یک مخزن و همچنین میزان مسمومیت آن را می توان با استفاده از ارگانیسم های فیتوپلانکتون - شاخص های saprobity تعیین کرد. مخازن پلی، مزو و الیگوساپروبیک وجود دارد. با توجه به سیستم کلاسیک موجودات شاخص ایجاد شده توسط کلکویتز و مارسون، Euglena viridis را می توان به عنوان polysaprobionts، Chlorella vulgaris به عنوان alpha-mesosaprobionts و Synura uvella به عنوان oligosaprobionts طبقه بندی کرد. از آنجایی که روشهای کنترل هیدروبیولوژیکی کیفیت آب به طور مداوم در حال بهبود است، فهرست موجودات شاخص صمیمیت در حال گسترش و اصلاح است.
افزایش اوتروفیکاسیون، یا تجمع بیش از حد مواد آلی در یک مخزن، ارتباط نزدیکی با افزایش فرآیندهای فتوسنتز در فیتوپلانکتون دارد. توسعه گسترده جلبک ها منجر به بدتر شدن کیفیت آب و "شکوفایی" آن می شود.
گلدهی یک پدیده خودبخودی نیست، در یک دوره زمانی نسبتا طولانی، گاهی اوقات دو یا چند فصل رشد آماده می شود. پیش نیاز افزایش شدید تعداد فیتوپلانکتون ها وجود جلبک ها در مخزن و توانایی آنها برای تولید مثل در شرایط مساعد است. به عنوان مثال، توسعه دیاتوم ها تا حد زیادی به محتوای آهن در آب بستگی دارد؛ عامل محدود کننده جلبک های سبز نیتروژن است و برای جلبک های سبز آبی - منگنز. اگر بیوماس فیتوپلانکتون در محدوده 0.5-0.9 میلی گرم در لیتر، متوسط - 1-9.9 میلی گرم در لیتر، شدید - 10-99.9 میلی گرم در لیتر باشد، در صورتی که زیست توده فیتوپلانکتون در محدوده 0.5-0.9 میلی گرم در لیتر باشد، شکوفه آب ضعیف در نظر گرفته می شود، و در صورت هایپرشکوفایی بیش از 100 میلی گرم در لیتر است. .
روش های مبارزه با این پدیده هنوز آنقدر پیشرفته نیست که بتوان مشکل را کاملاً حل شده در نظر گرفت. از اقدامات هیدرولوژیکی، پرکاربردترین و ایمن ترین آنها افزایش دبی و هوادهی مصنوعی مخزن است.
مشتقات کاربامید - دیورون و مونورون - به عنوان جلبک کش (عوامل شیمیایی برای مبارزه با گلدهی) در دوزهای 0.1-2 میلی گرم در لیتر استفاده می شود. برای تمیز کردن موقت مناطق جداگانه مخازن، سولفات آلومینیوم اضافه می شود. با این حال، باید با احتیاط به آفت کش ها متوسل شد، زیرا آنها نه تنها برای موجودات آبزی، بلکه برای انسان نیز خطرناک هستند.
در سال های اخیر، ماهی های گیاهخوار به طور گسترده ای برای این اهداف مورد استفاده قرار گرفته اند. بنابراین، کپور نقره ای انواع مختلفی از جلبک های پروتوکوکی، اوگلنوفیت و دیاتومه را مصرف می کند. سبزهای آبی که در طول رشد انبوه متابولیت های سمی تولید می کنند، به راحتی توسط آنها جذب می شوند، اما می توانند سهم قابل توجهی در رژیم غذایی نمونه های بالغ این ماهی را تشکیل دهند. فیتوپلانکتون ها نیز به آسانی توسط تیلاپیا، کپور نقره ای، کپور سر بزرگ، و در صورت کمبود غذای اساسی - ماهی سفید، بوفالو دهان بزرگ و ماهی پارویی خورده می شوند.
تا حدی ماکروفیت ها می توانند شدت شکوفه های آب را نیز محدود کنند. آنها علاوه بر رهاسازی مواد مضر برای فیتوپلانکتون ها در آب، سطح نواحی مجاور را سایه می اندازند و از فتوسنتز جلوگیری می کنند.
هنگام محاسبه منبع غذایی یک مخزن و تولید فیتوپلانکتون، لازم است ترکیب گونه، تعداد سلول و زیست توده جلبک بر اساس محتوای موجود در حجم معینی از آب (0.5 یا 1 لیتر) تعیین شود.
تکنیک پردازش نمونه شامل چندین مرحله (ثابت، تغلیظ، کاهش به حجم معین) است. تثبیت کننده های مختلفی وجود دارد، اما رایج ترین مورد استفاده فرمالین است (2-4 میلی لیتر محلول فرمالین 40 درصد در هر 100 میلی لیتر آب). به سلول های جلبک اجازه داده می شود تا دو هفته ته نشین شوند (اگر حجم نمونه کمتر از 1 لیتر باشد، دوره ته نشینی بر این اساس کوتاه می شود). سپس لایه بالایی آب ته نشین شده با دقت برداشته می شود و 30-80 میلی لیتر برای کار بیشتر باقی می ماند.
سلول های فیتوپلانکتون در بخش های کوچک (0.05 یا 0.1 میلی لیتر) شمارش می شوند، سپس محتوای آنها در 1 لیتر بر اساس نتایج به دست آمده تعیین می شود. اگر تعداد سلول های یک گونه جلبک خاص از 40 درصد تعداد کل آنها بیشتر شود، این گونه غالب در نظر گرفته می شود.
تعیین زیست توده فیتوپلانکتون یک فرآیند کار فشرده و زمان بر است. در عمل، برای تسهیل محاسبات، به طور معمول پذیرفته شده است که جرم 1 میلیون سلول فیتوپلانکتون آب شیرین تقریباً برابر با 1 میلی گرم است. روش های اکسپرس دیگری نیز وجود دارد. با توجه به نقش زیاد فیتوپلانکتون ها در اکوسیستم مخازن، در شکل گیری بهره وری ماهیان آنها، لازم است تمامی پرورش دهندگان ماهی، از دانشمندان گرفته تا متخصصان، به این روش ها تسلط داشته باشند.
اس. یودین
معرفی
فصل 1. مواد و روش تحقیق 7
فصل 2. ویژگی های منطقه مورد مطالعه 13
فصل 3. فیتوپلانکتون، پویایی فصلی و توزیع فضایی آن - 21
3.1 دینامیک فصلی ترکیب کیفی فیتوپلانکتون در جریانهای آب22
3.2. ویژگی های توزیع فضایی فیتوپلانکتون - 27
3.3. تجزیه و تحلیل خوشه ای از توزیع فضایی فیتوپلانکتون - 49
3.4. ارتباط بین توزیع فضایی فیتوپلانکتون و عوامل هیدرولوژیکی و پدیده رسوب گذاری فیتوپلانکتون در جبهه دلتا 52
فصل 4. دینامیک فصلی و توزیع فضایی رنگدانه های فتوسنتزی پلانکتون - 55
4.1 دینامیک فصلی کلروفیل a و سایر رنگدانه های فتوسنتزی پلانکتون در جریان های آب - 57
4.2. توزیع فضایی و زمانی کلروفیل a و سایر رنگدانه های فتوسنتزی پلانکتون در جبهه دلتا - 79
4.3. رابطه بین محتوای کلروفیل a و زیست توده فیتوپلانکتون - 88
فصل 5. تولید پلانکتون اولیه 92
5.1. دینامیک فصلی و توزیع فضایی 92
5.2. کارایی استفاده از انرژی تابش خورشیدی در طی فتوسنتز پلانکتون 98
5.3. تولید اولیه پلانکتون در دلتای ولگا در مقایسه با دهانه رودخانه های دیگر - 102
فصل 6. تحلیل گذشتهنگر شاخصهای بهرهوری فیتوپلانکتون و ارتباط آنها با عوامل مختلف - 109
6.1. تولید پلانکتون اولیه و وضعیت تغذیه ای ناحیه تحتانی دلتای ولگا در دوره های مختلف جریان تنظیم شده 109
6.2. رابطه بین شاخص های بهره وری فیتوپلانکتون و عوامل محیطی 115
6.4. تأثیر ورودی انسانزایی مواد مغذی و پارامترهای جریان آب ولگا بر بهرهوری فیتوپلانکتون دلتا 132
نتیجه گیری 138
ادبیات 140
معرفی کار
مرتبط بودن مشکل . یکی از وظایف اصلی هیدروبیولوژی ایجاد نظریه ای در مورد بهره وری بیولوژیکی اکوسیستم های آبی است (Alimov, 2001). با توجه به موضع پذیرفته شده G. G. Vinberg (1960)، تولید اولیه اکوسیستم های آبی، همراه با مواد آلی آلوکتونی که وارد آنها می شود، پایه مادی و انرژی تمام مراحل بعدی فرآیند تولید را تشکیل می دهد. مطالعات کمی در مورد شدت فرآیندهای تولید، در درجه اول تولید اولیه، اساس سیستم مدرن گونه شناسی مخازن است (Bouillon، 1983). تعیین شاخص های بهره وری فیتوپلانکتون بخشی جدایی ناپذیر از پایش کیفیت آب است (آباکوموف، سوشچنیا، 1992؛ اوکسیوک و همکاران، 1993).
دلتای ولگا یک شی طبیعی منحصر به فرد است که مهم ترین عملکرد زیست کره در حفظ هموستاز تالاب های منطقه را انجام می دهد و از اهمیت اقتصادی بالایی برخوردار است. در طول تقریباً نیم قرن گذشته جریان تنظیم شده ولگا، تغییرات قابل توجهی در شرایط محیطی در دلتای آن رخ داده است که عمدتاً به دلیل نوسانات در جریان آب، توزیع مجدد درون سالانه انسانی آن، تغییر سطوح آلاینده ها و مواد مغذی و سایر عوامل انسانی و طبیعی است. عوامل. در این راستا، تدوین مفهوم پایداری زیستمحیطی و آسیبپذیری اکوسیستمهای آبی منطقه و ارزیابی بهرهوری بیولوژیکی آنها از اهمیت ویژهای برخوردار است. بنابراین، مطالعه بهره وری فیتوپلانکتون در دلتای ولگا به ویژه در شرایط فعلی مرتبط است.
هدف و اهداف مطالعه . هدف از این کار بررسی بهره وری فیتوپلانکتون در دلتای ولگا است.
مطابق با هدف، وظایف زیر تدوین شد:
بررسی پویایی فصلی و توزیع فضایی فیتوپلانکتون.
مطالعه دینامیک فصلی و توزیع فضایی رنگدانه های گیاهی در پلانکتون.
بررسی پویایی فصلی و توزیع فضایی شدت فتوسنتز.
4. رابطه بین غلظت کلروفیل را تجزیه و تحلیل کنید آبا
زیست توده فیتوپلانکتون و شدت فتوسنتز.
شناسایی عوامل محیطی اصلی که بهرهوری فیتوپلانکتونها را تعیین میکنند.
وضعیت فعلی تولید پلانکتون اولیه را در چند سال جریان آب تنظیم شده ارزیابی کنید.
تازگی علمی . یک ارزیابی جامع از وضعیت فعلی بهرهوری فیتوپلانکتون و وضعیت تغذیهای پایین دستهای دلتای ولگا انجام شد. برای اولین بار، محتوا و دینامیک رنگدانه های گیاهی در پلانکتون به طور دقیق مورد مطالعه قرار گرفت، مقادیر عدد جذب به عنوان معیاری از فعالیت فتوسنتزی کلروفیل به دست آمد. آدر آب های مورد مطالعه ویژگیهای تولید اولیه پلانکتون در ناحیه تحتانی دلتا و جلو دلتا ولگا آشکار میشود و فرآیندهای حذف و ته نشینی فیتوپلانکتون در جبهه دلتا در نظر گرفته میشود.
اهمیت عملی . دلتای ولگا و منطقه ساحلی پیش از مصب آن نقش مهمی در حفظ تعادل اکولوژیکی در پهنه خشکی و پهنه آبی وسیع مجاور دریای خزر دارد. نتایج بهدستآمده میتواند مبنایی برای توسعه روشهایی برای استفاده منطقی و حفاظت از منابع آب بیولوژیکی باشد.
اشیاء در منطقه، برنامه ریزی اقداماتی برای افزایش بهره وری آنها، ساخت مدلی از تولید بیولوژیکی تحت گزینه های مختلف برای تنظیم جریان ولگا و تغییر جزء انسانی هجوم مواد مغذی. مواد تحقیقاتی به عنوان بخشی جدایی ناپذیر از سیستم نظارتی انجام شده توسط ذخیره گاه زیست کره آستاراخان گنجانده شده است و همچنین در فرآیند آموزشی موسسه زیست شناسی و مدیریت زیست محیطی ASTU استفاده می شود.
تایید کار . مطالب پایان نامه در کنفرانس علمی "وضعیت، مطالعه و حفاظت از مجموعه های طبیعی ذخیره گاه زیست کره آستاراخان در شرایط افزایش سطح دریای خزر و افزایش فشار انسانی" ارائه شد (آستراخان، 1999). کنفرانس بین المللی "فناوری های جدید در حفاظت از تنوع زیستی در اکوسیستم های آبی" (مسکو، 2002). در دومین کنفرانس بین المللی "بیوتکنولوژی - حفاظت از محیط زیست" (مسکو، 2004). کنفرانس بین المللی "مشکلات و چشم اندازهای احیای اکوسیستم های فناورانه" (آستراخان، 2004)؛ در کنفرانس بین المللی "تولید اولیه اکوسیستم های آبی" (Bo-rock, 2004); در کنفرانس های علمی و عملی کارکنان آموزشی دانشگاه فنی دولتی آستاراخان در سال 1999-2004.
انتشارات . 8 اثر با موضوع پایان نامه منتشر شده است.
ساختار و محدوده پایان نامه . پایان نامه در 163 صفحه، شامل 12 جدول، 26 شکل ارائه شده است. شامل یک مقدمه، 6 فصل و نتیجه گیری است. کتابشناسی شامل 230 عنوان است که 70 عنوان آن به زبان های خارجی است.
ویژگی های توزیع فضایی فیتوپلانکتون
در مسیرهای آب منطقه دلتای پایین و در جبهه دلتا، شرایط اکولوژیکی مختلفی ایجاد می شود که در ترکیب گونه ای فیتوپلانکتون، توزیع و فراوانی آن منعکس می شود. تفاوت در رشد جلبک ها را نیز می توان با توجه به فصول سال ردیابی کرد. در طول فصل رشد، چندین دوره را می توان با الگوی مشخص توزیع و رشد کمی فیتوپلانکتون ها متمایز کرد.
کمترین رشد کمی فیتوپلانکتون در همه انواع آب در فصل سرد مشاهده می شود. در همان زمان، موقعیت غالب در کانال ها توسط نمایندگان کلاس دیاتومه های پنتی، عمدتاً جنس Cymbella، Gomphonema، NavicuJa، Nitzschia، Synedra اشغال شده است. زیست توده کل فیتوپلانکتون معمولاً از 0.01-0.30 گرم مکعب تجاوز نمی کند.
در بهار، قبل از شروع سیل، با افزایش تابش خورشیدی و گرم شدن آب، آغاز رشد انبوه فیتوپلانکتون ها مشاهده می شود. آب به طور ناهموار گرم می شود و توزیع فضایی فیتوپلانکتون ها با لکه های بزرگ مشخص می شود. بیشترین گرمایش در ناحیه ساحلی جزایر آواندلتا مشاهده می شود. در این مناطق، زیست توده فیتوپلانکتونها از سایر مناطق بیشتر است و حدود 0.8-0.9 گرم در متر مربع و در برخی ایستگاهها بیش از 1 گرم در متر مربع است (جدول 3.2.1). توزیع فیتوپلانکتون در این دوره در شکل نشان داده شده است. 3.2.1.a. با استفاده از مثال آوریل 2000. دیاتومه های پنتی سهم بزرگی در زیست توده فیتوپلانکتون در نواحی ساحلی جزایر جلوی دلتا دارند که 50 تا 75 درصد از کل زیست توده را تشکیل می دهند. اول از همه، اینها نمایندگان جنس Surirella و Nitzschia هستند.
جلبکهای تقسیمبندی کلروفیتا (Chiorella، Pediastrum، Closterium) و Cyanophyta (Lyngbya، Oscillatoria) نیز در مقادیر قابلتوجهی رشد میکنند و در برخی از ایستگاهها موقعیت غالب را اشغال میکنند. در نواحی باز جبهه دلتا، رشد کمی فیتوپلانکتون در این دوره کمتر است، زیست توده حدود 0.3-0.4 گرم است، در کانال های این دوره، زیست توده فیتوپلانکتون در مقایسه با مناطق باز جبهه دلتا. ، در سالهای مختلف کمتر یا قابل مقایسه با آن به میزان 0.04-0.4 گرم"3 مشاهده می شود. گونه های غالب فیتوپلانکتون های زمستانی هستند که در درجه اول نمایندگان جنس های Niizschia و Cymbella و همچنین Aulacosira، Stephanodiscux، Amphora، Cocconeis، Gornphonema، Navicula، Rhoicosphenia هستند.
در نیمه دوم اردیبهشت ماه در اوج سیل، اوج توسعه فیتوپلانکتون در کانال ها وجود دارد که می توان آن را بهار و تابستان نامید. در عین حال، زیست توده فیتوپلانکتون دارای توزیع فضایی نسبتاً یکنواختی است و ترکیب گونهای مشابه مشاهده میشود. این امر با این واقعیت توضیح داده می شود که در شرایط سیلاب، سطح آب بالا و سرعت جریان بالا، از یک سو، شرایط یکسانی را به ویژه در کانال ها و مناطق باز جبهه دلتا تعیین می کند و از سوی دیگر، ایجاد می کند. رانش ترانزیت قابل توجه جلبک های پلانکتون توزیع فیتوپلانکتون در اوج سیل در شکل 1 نشان داده شده است. 3.2L.6. با استفاده از مثال می 2000. در این دوره، دیاتومه های مرکزی نقش اصلی را در کانال های میان بخش Bacillariophyta به دست می آورند، در درجه اول به دلیل پوشش گیاهی انبوه Stephanodiscvs hantzschii. جلبک های بخش کلروفیتا به ویژه Chiorella vulgaris Beijer به طور قابل توجهی در حال توسعه هستند.این گونه ها (S. hantzschli و Ch. vulgaris) به مقدار قابل توجهی در اکثر ایستگاه ها در مناطق باز و ساحلی جبهه دلتا یافت می شوند. زیست توده فیتوپلانکتون در کانال ها به 0.5-1.8 گرم مکعب می رسد، در مناطق باز جبهه دلتا - 0.5-0.8 گرم، در مناطق ساحلی جزایر جلوی دلتا - 0.1-0.3 گرم می رسد. فیتوپلانکتون اوج توسعه در طول دوره سیل جایگزین می شود. با کاهش قابل توجهی که اغلب در نیمه دوم ژوئن رخ می دهد،
با استقرار کم آب تابستان-پاییز، بیشترین تمایز در توزیع فضایی فیتوپلانکتون مشاهده می شود. توزیع فیتوپلانکتون در این دوره در شکل نشان داده شده است. 3.2.1، با استفاده از مثال آگوست 2000. در کانال ها، زیست توده فیتوپلانکتون افزایش می یابد، در حالی که تعداد گونه ها کاهش می یابد. به طور معمول، پویایی رشد فیتوپلانکتون ها در طول دوره کم آب تابستان-پاییز دارای ویژگی چند اوج است؛ در طی پیک ها، زیست توده جلبک در برخی سال ها به 2.0-4.0 گرم در مترمکعب افزایش می یابد و به مقادیر بالاتر می رسد. بنابراین در سال 1997 در پایان در ماه جولای، زیست توده فیتوپلانکتون به 16.5 گرم رسید. جلبک های بخش Baclariophyta غالب هستند که زیست توده آنها معمولاً حدود 85-95٪ از ارزش کل است. در این مورد، به دلیل توسعه جلبکهای جنس Aulacosira (A. gmnulata f. granulata، Aulacosira sp.) و Sceleionema (S. subsalsum) سهم اصلی بر دیاتومههای مرکزی است. همچنین نیمه دوم تابستان فصل رشد جلبکهای سبز آبی، عمدتاً Aphanizomenon flos-aqua و Microcystis aeruginosa است که در برخی سالها رشد کمی آنها میتواند قابل توجه باشد. در مناطق باز آواندلتا، زیست توده فیتوپلانکتون تا پایان ماه اوت به 0.03-0.3 گرم کاهش می یابد.» در عین حال، سهم باکلاریوفیتا در زیست توده کل فیتوپلانکتون به دلیل کاهش زیست توده جلبک ها به طور قابل توجهی افزایش می یابد. ارزش زیست توده فیتوپلانکتون در ایستگاه های مختلف بخش های ساحلی جزایر آواندلتا به طور قابل توجهی متفاوت است که به دلیل تنوع بیوتوپ ها و تأثیر عوامل محلی است. در اکثر ایستگاه ها مقادیر بالایی مشاهده می شود - در محدوده 0.5-4.5 hm3، در برخی موارد به مقادیر بالاتر می رسد.بیشترین بیومس مشاهده شده فیتوپلانکتون ساحلی 27.5 ru J بود. توزیع ترکیب گونه ها موزاییک است.
ارتباط بین توزیع فضایی فیتوپلانکتون ها و عوامل هیدرولوژیکی و پدیده رسوب گذاری فیتوپلانکتون در جبهه دلتا
با برهمکنش دینامیکی رودخانه و مخزن دریافت کننده در داخل منطقه مصب رودخانه، آب رودخانه گسترش یافته و نرخ جریان در ساحل کاهش می یابد (River Deltas, 1986). در ناحیه کم عمق خور ولگا در نزدیکی ساحل، با یک جریان مسطح، سرعت جریان ها در ابتدا به شدت در دهانه آبراهه در لبه دریای دلتا (در دهانه رودخانه "میکروبار") کاهش می یابد، و سپس تقریباً تا زمانی که آب از نوار دریا خارج نشود تغییر نکنید (Ustevaya.,., 1998). در کانال ها، سرعت جریان در طول دوره سیلاب 0.51-1.30 ms"1، در دوره کم آب - 0.08-0.36 ms"1، در جبهه دلتا این ارقام به ترتیب 0.25-0.62 ms"1 و 0.04-0.12 می باشد. m "s"[ (Moskalenko، 1965). چنین تغییر قابل توجهی در نرخ جریان در هنگام حرکت از کانال ها به جبهه دلتا یکی از عوامل اصلی تعیین کننده تغییر در ترکیب و فراوانی فیتوپلانکتون است. با استقرار کم آب تابستان-اوسپی، با حرکت از دهانه کانال ها به سمت نواحی باز جبهه دلتا به دلیل تخریب مجموعه فیتوپلانکتون رودخانه، که هسته آن در طول این مدت است، کاهش این شاخص ها مشاهده می شود. دوره در درجه اول نمایندگان جنس Aulacosira و Melosira است.
تعداد زیادی کار وجود دارد که اشاره می کند که برای توسعه بسیاری از دیاتوم ها، جریان و اختلاط آشفته ضروری است و برای حفظ حالت معلق از اهمیت بالایی برخوردار است (Lund، 1966؛ Oksiyuk، 1973؛ Kiselev، 1980؛ Oksiyuk، Stolberg، 1988؛ هیدروبیولوژی. ...، 1990، و غیره). به گفته K.A. Guseva (Guseva، 1968)، بسیاری از دیاتومه های پلانکتونیک آب شیرین، به ویژه جنس Melosira، سازگاری قابل اعتمادی برای اوج گرفتن ندارند، بنابراین به حرکت مداوم آب نیاز دارند.
ما با توجه به کاهش کلروفیل a در پلانکتون در طول زمان در طول زمان نشست، آزمایشهای آزمایشگاهی را در استوانههای پر از آب گرفتهشده از یک کانال در طول دوره کمآب تابستان-پاییز انجام دادیم. ترکیب فیتوپلانکتون ها برای این فصل معمولی بود - سهم اصلی را دیاتومه های مرکزی تشکیل می دادند، در درجه اول جنس Aulacosira و Melosira (93٪). مقادیر به دست آمده در شرایط آزمایشگاهی به ارتفاع لایه برابر با عمق متوسط جبهه دلتا کاهش یافت. منحنی تجربی به دست آمده از سینتیک از دست دادن کلروفیل a در طول ته نشینی با منحنی کاهش کلروفیل a در جبهه دلتا با حرکت از دهانه کانال ها به سمت دریا مقایسه شد (شکل 3.4.1). برای مقایسه نتایج، دادههای مربوط به محتوای کلروفیل a در جبهه دلتا با در نظر گرفتن فاصله ایستگاهها از دهانه کانالها و زمان عبور تودههای آب از این فاصله ارائه شد.
همانطور که در شکل 3.4.1 مشاهده می شود، ماهیت منحنی کاهش کلروفیل a در پلانکتون در آزمایش و در شرایط طبیعی مشابه است، که نشان دهنده نقش مهم عامل رسوب گرانشی فیتوپلانکتون در جبهه دلتا است. در عین حال، کاهش کلروفیل a در آزمایش کندتر از جبهه دلتا رخ می دهد. اول از همه، این را می توان علاوه بر رسوب گرانشی فیتوپلانکتون در جبهه دلتا، با فیلتر کردن و مصرف آن توسط موجودات بنتوس و پری فوتون توضیح داد.
بنابراین، در قسمت پایین دلتای ولگا، موجودات فیتوپلانکتون، همراه با توده های آب، از کانال ها به سمت جبهه دلتا منتقل می شوند. در اینجا یک تغییر سریع در ترکیب گونهای فیتوپلانکتون و تبدیل مجتمعهای پلانکتون رودخانه به دلتا-جلوها وجود دارد. در همان زمان، اشکال رودخانه های معمولی از ترکیب پلانکتون ناپدید می شوند یا تعداد آنها کاهش می یابد. زیست توده فیتوپلانکتون کاهش می یابد، محتوای کلروفیل a در پلانکتون ها و شدت فتوسنتز کاهش می یابد، زیرا در شرایط آب کم عمق، رشد بیش از حد شدید پوشش گیاهی آبزی و کاهش شدید جریان، فرونشست و مرگ رخ می دهد.
شناخته شده است که موجودات پلانکتون، به ویژه فیتوپلانکتون، متمرکزکننده های خاص بسیاری از عناصر شیمیایی هستند؛ آنها نقش مهمی در مهاجرت بیوژنیک آنها دارند (Telitchenko و همکاران، 1970؛ Khobotyev، Kapkov، 1972؛ Varenko، Misyura، 1985). ته نشین شدن فیتوپلانکتون رودخانه در جبهه دلتا باید منجر به آزاد شدن برخی از عناصر شیمیایی که قبلاً بخشی از موجودات پلانکتون بوده یا ورود آنها به خاک همراه با ته نشین شدن پلانکتون ها باشد.
اخیراً ارزیابی بهرهوری اولیه بدنههای آبی علاوه بر روشهای سنتی، با تعیین ارزش کلروفیل a در پلانکتونها انجام شده است. این روش برای نظارت بر محیط زیست در همه انواع آب - اعم از آب شیرین و دریایی استفاده می شود. شاخص های محتوای کلروفیل a برای تعیین وضعیت تغذیه ای بدنه های آبی استفاده می شود (Vinberg، 1960؛ Pyrina، 1965a؛ Mineeva، 1979، و غیره). غلظت کلروفیل a برای تعیین زیست توده جلبک ها استفاده می شود (Vinberg, 1960; Elizarova, 1975, 1993). دادههای مربوط به محتوای کلروفیل a در پلانکتون برای تعیین تولید اولیه غیر ارگانیک فیتوپلانکتون و مدلسازی این فرآیندها به کار میرود (Pyrina، Elizarova، Nikolaev! 973؛ Makarova، Zaika، 1981، و غیره). برای توصیف کیفیت آب، از نسبت شفافیت کلروفیل به آب استفاده می شود (Bulyok، 1977؛ 1983). انباشت کلروفیل a در حجم آب برای ارزیابی درجه اوتروفیکاسیون بدنههای آبی و وضعیت بهداشتی و بیولوژیکی آنها عمل میکند.» همبستگی مثبتی بین توزیع کلروفیل a توسط کاروتنوئیدها و فتوسنتز یافت شد. بر اساس نسبت کلروفیل a و کاروتنوئیدها، وضعیت فیزیولوژیکی جمعیت جلبک ها و وضعیت محیط را می توان ارزیابی کرد (Margalef, I960; 1967؛ Watson, Osborne, 1979؛ Davydova, 1983).
بنابراین، ارزش محتوای کلروفیل a در آب یک ویژگی جدایی ناپذیر از بهره وری بیولوژیکی بدنه های آبی در سطح اتوتروف است که یک شاخص مهم از وضعیت اکوسیستم های آبی است.
توزیع فضایی و زمانی کلروفیل a و سایر رنگدانه های فتوسنتزی پلانکتون در جبهه دلتا
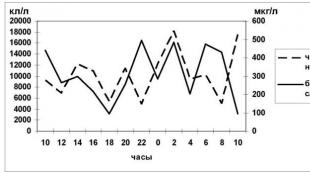
توزیع کلروفیل a و سایر رنگدانه های فتوسنتزی پلانکتون در ناحیه آبی جبهه دلتا با یک الگوی موزاییک بزرگ مشخص می شود. تغییرات فصلی در توزیع فضایی کلروفیل a، علیرغم برخی تفاوت ها در سال های مختلف، عموماً با ماهیت کلی دینامیک مشخص می شود. دینامیک فصلی محتوای رنگدانه های فیتوپلانکتون در مناطق مختلف جبهه دلتا در جدول ارائه شده است. 4.2.1،4-2.2، 4-2.3،4،2-4 در بهار، قبل از شروع سیل، در جبهه دلتا، غلظت کلروفیل a در سال های مختلف در محدوده 4.7-91.7 میلی گرم در متر بود. در اکثر ایستگاهها، محتوای کلروفیل a نسبتاً یکنواخت بود و از 9.6 تا 12.7 میلیگرم در متر متغیر بود. غلظت های بالاتر کلروفیل a در مناطق ساحلی جزایر جلوی دلتا - تا 31.2 میلی گرم در متر "و در مناطق محلی در مناطق ساحلی جزایر با کاهش تبادل آب - تا 91.7 میلی گرم در متر مشاهده شد. در کانالهای موجود در این زمان، مقدار کلروفیل a به طور قابل توجهی کمتر بود و به میزان 2.3 - 3.5 میلی گرم در متر مکعب بود. دلتای بالای آب در فصل بهار به دلیل رشد زودتر فیتوپلانکتون ها در جبهه دلتا در شرایط گرم شدن بهتر و افزایش دمای آب است، اختلاف دمای آب در کانال ها و جبهه دلتا در این دوره می تواند به بیش از 15 درجه - در کانال ها دمای آب از 8.0-12.0 C تجاوز نمی کند، در جبهه دلتا آب می تواند تا 17.5-20،0C در مناطق باز و تا 25.0-27.5C در مناطق ساحلی جزایر گرم شود. محتوای رنگدانه های اضافی در این دوره کم بود.غلظت کلروفیل b به طور متوسط 7-14% کلروفیل کلروفیل کلروفیل c - 11-16% بود. غلظت کاروتنوئیدها بین 6.1 تا 80.9 mSPU-m بود. و در اکثریت قریب به اتفاق ایستگاه های بررسی شده اندکی از غلظت کلروفیل O فراتر رفته است. مقادیر نسبت رنگدانه مشاهده شده در محدوده نسبتاً گسترده ای متفاوت بود - از 0.8 تا 1.6، اما در اکثر ایستگاه های بررسی شده پایین بود.
در طول دوره سیلاب بهار و تابستان، غلظت کلروفیل a در جبهه دلتا در محدوده 1D-22.9 mg-m"3، مقادیر متوسط 5.8-14.7 mg-m"3 بود. بیشترین مقادیر کلروفیل a در نواحی باز جبهه دلتا مشاهده شد که عمدتاً به دلیل حذف پلانکتون از کانال ها بود که با سرعت جریان بالا تسهیل شد. در کانال ها در این زمان، افزایش غلظت کلروفیل a به 9.B-mgm - 22.8 MG G\ مشاهده شد که کمترین مقدار در مناطق ساحلی جزایر مشاهده شد، جایی که در مقایسه با قبلی در دوره، غلظت کلروفیل a 2-6 برابر کاهش یافت به طور کلی، توزیع کلروفیل a در دلتای جلویی در طول دوره سیل بهار و تابستان با بیشترین یکنواختی مشخص می شود.
میزان کلروفیلهای d و c نسبت به کل کلروفیلها در جبهه دلتا به ترتیب از 0 تا 14.1 درصد و O تا 18.3 درصد بود. مناطق جلوی دلتا باز. غلظت کاروتنوئیدها از 079 تا 21.0 mSPU-m"3 بود و به طور کلی از نظر مقدار نزدیک به غلظت کلروفیل a بود.
در طول کاهش سیل، غلظت کلروفیل a در مناطق باز جبهه دلتا کاهش یافت، در نواحی ساحلی جزایر کمی تغییر کرد.در دوره کم آب (تیر، مرداد) غلظت کلروفیل a به طور کلی کم بود، میانگین غلظت 3.6-6.3 میلی گرم در متر مکعب بود.در مناطق محلی در نواحی ساحلی جزایر، غلظت کلروفیل در حدود قابل توجهی متغیر بود و می توانست به مقادیر بسیار بالایی برسد (تا 2/97 میلی گرم بر متر). مقدار کلروفیل بیس از 0 تا 27.8 میلی گرم در متر مربع و از 0 تا 29.4 میلی گرم در مترمربع متغیر بود. سهم کلروفیل b از کل مقدار کلروفیل 0 - 19D، کلروفیل c - 0- بود. 21.7٪ غلظت کاروتنوئیدها به طور متوسط 4.4-38.4 mSPU-m" و در اکثر ایستگاه ها از غلظت کلروفیل a فراتر رفت. نسبت رنگدانه بین 0.8 - 2.0 متغیر بود، بالاترین مقادیر در جبهه دلتای باز مشاهده شد، جایی که در در آن زمان مانعی در رشد فیتوپلانکتون وجود داشت. در کانال ها در این دوره، غلظت کلروفیل a به طور کلی بیشتر بود.
در پاییز، در ماه های سپتامبر و اکتبر، در اکثر مناطق آبی دلتای پیشین، میزان کلروفیل a در محدوده 1.5-13.3 میلی گرم در متر مربع 9 و در نواحی ساحلی جزایر به طور قابل توجهی بود. بالاتر - 9.2-113.2 mg-m "3. تصویر کلی از توزیع فضایی رنگدانه های گیاهی به طور کلی شبیه به آنچه در پایان تابستان مشاهده شد بود.
توزیع فضایی کلروفیل a در شکل 1 نشان داده شده است. 4.1 L. (با استفاده از مثال 2000).
بنابراین، توزیع فضایی رنگدانههای فتوسنتزی پلانکتون در جبهه دلتا با ناهمواری زیاد مشخص میشد و با فصول متفاوت بود.
بیشترین مقدار کلروفیل a در جبهه دلتا در فصل بهار، قبل از شروع سیل، به ویژه در نواحی ساحلی جزایر مشاهده شد، زمانی که با افزایش تابش خورشیدی در شرایط آب کم عمق، گرم شدن قابل توجهی در سطح آب مشاهده شد. آب به طور قابل توجهی بیشتر از کانال ها رخ می دهد.
با شروع آب زیاد، غلظت کلروفیل a در نواحی ساحلی جزایر کاهش یافت و در جبهه دلتای باز کمی تغییر کرد. در این دوره، یکنواختترین توزیع فضایی کلروفیل a در کانالهای دلتای بالای آب و در آواندلت مشاهده شد، زیرا از یک سو سطح آب بالا و سرعت جریان بالا به طور نسبی فراهم میشود. شرایط یکنواخت، و از طرف دیگر، تخریب ترانزیت فیتوپلانکتون رخ می دهد. در طول دوره تابستان و پاییز کم آب در جبهه دلتا، محتوای کلروفیل a به طور کلی کم است، به استثنای مناطق محلی مناطق ساحلی جزایر.
کارایی استفاده از انرژی تابش خورشیدی در طی فتوسنتز پلانکتون
مقدار انرژی تابش خورشیدی که در واحد سطح یک مخزن افت می کند، که در نتیجه فعالیت فتوسنتزی پلانکتون در ستون آب زیر این ناحیه انباشته شده است، به صورت درصد بیان می شود، کارایی استفاده از انرژی تابش خورشید توسط پلانکتون ها را مشخص می کند. این شاخص به خوبی میزان توسعه فیتوپلانکتون ها را نشان می دهد و میزان تشکیل جدید مواد آلی را نشان می دهد. بر خلاف ارزش تولید اولیه، شاخص کارایی استفاده از انرژی تابش خورشیدی به روشنایی بستگی ندارد، که در درجه اول توسط شرایط آب و هوایی در طول دوره آزمایش تعیین می شود و اغلب هنگام مقایسه نتایج یک مانع است. بنابراین، کارایی استفاده از نور توسط پلانکتون ها اغلب می تواند شاخص مناسب تری برای بهره وری بدنه های آبی نسبت به ارزش واقعی تولید اولیه باشد (پیرینا، 1967).
مسئله کارآیی استفاده از انرژی نور خورشید در طول فتوسنتز پلانکتون به طور مفصل توسط G.G. وینبرگ (I960)، که خاطرنشان کرد که حداکثر مقادیر استفاده از انرژی خورشیدی توسط اسوالد (Oswald et al., 1957؛ به نقل از وینبرگ، 1960) در شرایط ویژه "برکه های اکسید کننده" به دست آمده است، جایی که میانگین آنها 2-4 است. درصد کل انرژی تابش فصل در مخازن طبیعی این مقادیر بسیار کمتر است. به عنوان نمونه ای از درجه بالای استفاده از انرژی خورشیدی، G.G. وینبرگ بازده استفاده از تشعشعات را برای دریاچههای چرنوی و کوسینو (طبق دادههای خود در سال 1937) در سطح 0.4 درصد در سال و 0.77 درصد - حداکثر در روز و برای دریاچه Zöllerod - 0.77 درصد حداکثر در روز ذکر میکند (Stccraann Nielsen, 1955؛ به نقل از وینبرگ، 1960). نتایج بالایی توسط I.L. Pyrina (1967) برای مخازن ولگا (ایوانکوفسکی، ریبیسکی و کویبیشفسکی) - به طور متوسط برای فصل رشد 0.2-0.5٪ و حداکثر روزانه 1.17-2.14٪ انرژی تابش مرئی. اگر در نظر بگیریم که نویسندگان پذیرفته اند که تابش مرئی 50٪ از کل را تشکیل می دهد، این مقادیر به ترتیب OD-0.25٪ و 0.58-1.07٪ از کل تابش خورشیدی خواهد بود. در مخزن Sheksninsky، متوسط راندمان استفاده از انرژی خورشیدی در طول فصل رشد زیر 0E08-0D 1٪ بود (Mineeva، 2003). به عنوان مثالی از راندمان پایین استفاده از انرژی تابش خورشیدی، G.G. وینبرگ دریاچه بلو را ذکر می کند، جایی که این مقدار از 0.04٪ تجاوز نمی کند و برای سال 0.02٪ از انرژی کل تابش را تشکیل می دهد. مقادیر کمتری نیز توسط L.G. کورنووی و ن.م. Mshieeva (1986) هنگام مطالعه مخازن با کدورت بالا. حداکثر مقادیری که آنها برای بازده استفاده از انرژی خورشیدی برای مخازن Vytegorsky و Novinkinsky، رودخانه Kovzha در دهانه و نزدیک روستای Annensky Most به دست آوردند، به ترتیب 0.06٪ بود. 0.02%؛ 0.05٪ و 0.007٪ و حداقل ها 0.02٪، 0.001٪، 0.006٪ و 0.001٪ هستند.
طبق دادههای ما، در جریانهای آب پایین دست دلتای ولگا، راندمان استفاده از انرژی تابش خورشیدی در طول فصل رشد به طور متوسط 0.04-0.06٪ از کل یا 0.10-0.12٪ تابش فعال فتوسنتزی است. راندمان استفاده از انرژی خورشیدی توسط فیتوپلانکتونها بر اساس فصل بسیار متفاوت بود (جدول 5.2.1.) بالاترین مقادیر در اوج توسعه فیتوپلانکتونها در ماه مه و ژوئیه-آگوست مشاهده شد، زمانی که میانگین بازده ماهانه استفاده از کل تابش خورشیدی بود. انرژی به 0.05-0.08٪ رسیده است مقادیر کم راندمان استفاده از انرژی خورشیدی مشخصه دوره های شدت کم فتوسنتز فیتوپلانکتون است - اوایل بهار و اواخر پاییز، زمانی که راندمان استفاده از کل انرژی تابش خورشیدی معمولاً تجاوز نمی کند. 0.03-0.04٪. در ماه ژوئن، با کاهش تولید اولیه پلانکتون، بازده نسبتاً پایینی در استفاده از کل انرژی تابش خورشیدی نیز مشاهده شد که ظاهراً با کمبود مواد مغذی در طول کاهش سیل همراه است.
بنابراین، میتوان نتیجه گرفت که پایین دستهای دلتای ولگا با مقادیر نسبتاً پایین بهرهوری انرژی تابش خورشیدی در طول فتوسنتز پلانکتون مشخص میشوند. راندمان پایین استفاده از انرژی خورشیدی توسط پلانکتون ها در کانال ها تا حد زیادی به دلیل محتوای بالای ذرات معلق در آب است که شفافیت کم آب ایجاد می کند و باعث تضعیف قابل توجه تشعشعات نافذ با عمق می شود. وابستگی درجه استفاده از نور در یک مخزن به شفافیت آب توسط Comita و Edmondson (1953) مورد توجه قرار گرفت. حرکت متلاطم توده های آب در کانال ها نیز از اهمیت بالایی برخوردار است که باعث ایجاد اختلاط مداوم می شود. در نتیجه یک یا قسمت دیگری از سلول ها همیشه جلبک ها به فراتر از منطقه یوفوتیک منتقل می شوند در حالت به اصطلاح "گرسنگی نور" در تاریکی باقی می مانند (Sorokin, 1958).
یک رابطه نزدیک بین شاخص های ماهانه بهره وری از انرژی تابش خورشیدی و غلظت کلروفیل "a" در پلانکتون کانال مشاهده شد (شکل 5.2.1.).
جداول 6.7-6.9 نتایج اندازه گیری های تولید اولیه در آب های صخره های مختلف را خلاصه می کند. همانطور که از جداول مشاهده می شود، تولید اولیه حتی در آب های همان صخره بسیار متفاوت است. با این حال، این نوسانات اخیر کاملا طبیعی هستند و در درجه اول با تغییرات در غلظت فیتوپلانکتون در بخش هایی از مناطق کم عمق صخره به سمت مرکز تالاب مرتبط هستند.
در تالاب های مرجانی با تولید اولیه کم (جزایر مرجانی جزیره لاو و جزایر مارشال، جزایر تواموتو)، حد پایین (4-2 میلی گرم C/m3 در روز) به مقادیر آن در الیگوتروفیک نزدیک است. آبهای گرمسیری جریانهای باد تجاری آنها را میشویند، جایی که معمولاً 1-5 میلیگرم C/m3 در روز است (Sorokin, 1971a). با این حال، حتی در چنین جزایر مرجانی، شیوع فصلی رشد فیتوپلانکتون ها در آب های تالاب های آنها ممکن است، که می تواند به شدت "شکوفه" برسد (Sournia, Ricard, 1976a). همانطور که از داده های جدول مشاهده می شود. 6.7، در تالابهای مرجانی، حتی آنهایی که توسط آبهای استوایی الیگوتروف با سطوح بسیار کم رشد فیتوپلانکتونها شسته میشوند، تولید اولیه اغلب با میانگین و گاهی سطح بالایی بهرهوری فیتوپلانکتون در حوضههای دریایی مزوتروف (15-50 میلیگرم سانتیگراد بر متر مکعب در هر متر مربع) مطابقت دارد. روز). در تالاب صخره حلقوی بزرگ Cargados-Carajos، واقع در هسته آبهای بسیار غیرمولد جریان باد تجاری جنوبی اقیانوس هند، تولید اولیه با زیست توده جلبکی (عمدتاً دیاتوم ها) به 250 میلی گرم در متر مکعب در روز رسید. ) بیش از 5.2 گرم بر متر مکعب (جدول 6.10). تولید بالا (70-100 میلی گرم C/m3 در روز) در تالاب بسته فانینگ آتول (جزایر لاو) در طول دوره توسعه داینوفلاژلات ها و جلبک های سبز آبی کوکوئید در آنجا ثبت شد (Gordon et al., 1971). این جزیره مرجانی نیز در آبهای الیگوتروف جریان باد تجاری قرار دارد. بنابراین، حتی در تالابهای صخرههای حلقهای منفرد واقع در جریان آبهای الیگوتروف، جوامع فیتوپلانکتون کاملاً تولیدی میتوانند وجود داشته باشند که تولید و زیست توده آنها 1-2 مرتبه بزرگتر از آبهای اقیانوسی است که آنها را میشویند.
با قضاوت بر اساس نتایج چند اندازه گیری موجود، غلظت کلروفیل a فعال فتوسنتزی در آب های تالاب صخره های حلقه ای اقیانوس باز به طور متوسط 5-10 برابر بیشتر از آب های اقیانوس اطراف است. با توجه به اندازه گیری های منفرد، در تالاب های صخره های مختلف در محدوده 0.2 تا 1 میلی گرم بر متر مکعب متغیر بود. در تالاب تاکاپوتو آتول، با توجه به نتایج مشاهدات طولانی مدت (Sougnia, Ricard, 1976a)، غلظت کلروفیل از 0.26 تا 0.77 میلی گرم بر متر مکعب متغیر بوده و در نقاط مختلف تالاب بسته به شدت ورود به آب های اقیانوس در طول جزر و مد.
سهم قابل توجهی در مطالعه فیتوپلانکتون ها و محصولات آن توسط مشاهدات فصلی انجام شده در آب های سد و صخره های ساحلی در نزدیکی جزایر بزرگ انجام شد، جایی که ایستگاه های بیولوژیکی برای مطالعه زندگی صخره ها و منطقه نریتیک مجاور اقیانوس ایجاد شد. در میان چنین صخرههایی که به خوبی مطالعه شدهاند، باید به صخرههای جزیره نوسی بی و تولئار (ماداگاسکار)، صخرههای هاوایی در جزیره اوآهو، صخرههای سد بزرگ استرالیا، صخرههای Waprao، Tiaura و Faaa در جزایر تاهیتی و موریا، صخرههای مرکزی ویتنام در نزدیکی Nha Trang. نتایج اندازه گیری تولید اولیه و زیست توده فیتوپلانکتون سدی و صخره های ساحلی در جدول خلاصه شده است. 6.8، 6.9، 6.10: داده های ارائه شده در آنها تقریباً برای همه صخره های مورد مطالعه بسیار متفاوت است. آنها منعکس کننده اندازه گیری های منظم تولید اولیه در مناطق مختلف یک صخره هستند. در عین حال، کوچکترین مقادیر آن (و همچنین کوچکترین مقادیر زیست توده فیتوپلانکتون: شکل 6.10) مربوط به مناطق کم عمق صخره است، جایی که آب در تماس شدید با جمعیت کف و فلور جلبکی است. از صخره در این مناطق (مسطح، صخره های تکه ای، ساحلی) فیتوپلانکتون، به عنوان یک قاعده، تخلیه می شود و تولید آن در اینجا به شدت کاهش می یابد. حداکثر مقادیر تولید فیتوپلانکتون و زیست توده ارائه شده در این جداول در هر صخره مربوط به مناطق تالاب یا شیب بیرونی حاشیه یا صخره ساحلی با عمق بیش از 15-20 متر است، جایی که آب در آن نیست. تماس مستقیم با جمعیت پایین صخره. ارزش تولید اولیه در این مناطق، به طور معمول، به طور قابل توجهی بالاتر از آب های دریای آزاد مجاور است. به طور متوسط، آنها بالاتر از آب تالاب های جزایر مرجانی و صخره های حلقوی اقیانوس باز هستند.
مقادیر بالای تولید اولیه، رسیدن به سطح آبهای اوتروفیک، در آب تعدادی از صخرههای ساحلی و مانع اقیانوس هند، مانند صخرههای ماداگاسکار، جزایر سیشل و جزیره سوکوترا و همچنین مشاهده شد. در تالاب صخره سد B. در طول دوره های "شکوفایی" جلبک های سبز آبی (نگاه کنید به شکل 6.9). تولید روزانه در اینجا به 50-300 میلی گرم C/m 3 در روز در لایه بالایی آب یا 0.7-2.8 گرم در متر مکعب رسید. علاوه بر این، در آب های صخره های سد جزایر شسته شده توسط آب های اقیانوس الیگوتروف (Tangi، Oahu، Lizard)، تولید اولیه در تالاب ها می تواند به 50-60 میلی گرم C/m2 یا 0.4-0.7 گرم کربن در هر 1 متر در روز برسد. که مربوط به سطح بالای بهره وری آبهای مزوتروفی است. در نواحی صخره ای سواحل ویتنام مرکزی و جزایر آن در دریای چین جنوبی (جزایر Thu) و در خلیج تایلند (جزایر Nam Zu) (به جدول 6.8، شکل 6.10 مراجعه کنید)، مقادیر فیتوپلانکتون اولیه تولید بالا بود، علیرغم این واقعیت که مشاهدات در طول دوره حداقل فصلی فیتوپلانکتون انجام شد (Sorokin et al., 1982). بالاترین مقادیر تولید اولیه، به عنوان یک قاعده، در آب های لبه بیرونی صخره در فاصله 10-100 متر از آن در بالای عمق 5-15 متر مشاهده شد (شکل 6.10 را ببینید). معمولاً بیش از 30-40 میلی گرم C/m 3 در روز بود. در سال 1984 در بخش های جداگانه این منطقه، تولید اولیه بالای 100 میلی گرم C/m3 در روز ثبت شد.
اطلاعات در مورد زیست توده فیتوپلانکتون ها در صخره های ساحلی و سدی هنوز بسیار پراکنده است. دقیق ترین اندازه گیری زیست توده فیتوپلانکتون در طول فصل توسط Angot (1968) در جزیره Nosy Be (ماداگاسکار) انجام شد. نتایج این اندازه گیری ها در شکل 1 خلاصه شده است. 6.9. روی میز جدول 6.10 داده های موجود در ادبیات مربوط به زیست توده فیتوپلانکتون در تعدادی از صخره های بزرگ اقیانوس های هند و اقیانوس آرام را ارائه می دهد. همانطور که از جدول مشاهده می شود، زیست توده فیتوپلانکتون در آب های سد و صخره های ساحلی می تواند به 3-8 گرم در متر مکعب برسد. کمترین مقدار آن در آب صخرههای ساحلی و سدی بخش مرکزی اقیانوس آرام مشاهده شد که در حالت افسرده قرار داشتند (صخرههای جزایر پانارورا و پااما، نیوهبریدها، صخرههای جزیره توووتا، جزایر لائو). ).
اطلاعات در مورد محتوای کلروفیل a در آب های سد و صخره های ساحلی به همان اندازه محدود است (جدول 6.11). دقیق ترین اندازه گیری های آن در آب های تالاب های جزایر مورآ و تاهیتی توسط محققان فرانسوی انجام شد (Ricard, 1976; Sournia, Ricard, 1976). این اندازه گیری ها نشان داد که غلظت کلروفیل در آنها در فصول مختلف سال در محدوده 0.03-0.17 میلی گرم بر متر مکعب متغیر است. در تالاب صخره هاوایی در خارج از جزیره اوآهو، و همچنین در تالاب صخره سد B. در زمستان، غلظت کلروفیل نزدیک به آن در آب های دریای مزوتروف بود: 0.1-0.3 میلی گرم بر متر مکعب. در طول دوره پاییز و تابستان حداکثر رشد فیتوپلانکتون در تالاب صخره سد B. و همچنین در آبهای صخره خارج از جزیره Nosy Be (ماداگاسکار)، غلظت کلروفیل در آب بیشتر بود. 1 میلی گرم بر متر مکعب، که نزدیک به محتوای آن در آب های اوتروفیک است (Angot، 1968). تأثیر تعلیق مجدد کل لایه رسوبات پایین، غنی شده با میکروبنتوزها، در ناحیه موج شکن بر غلظت کلروفیل در آبهای صخره ثابت شده است (Walker, 1981).
همانطور که در بالا ذکر شد، ارزش تولید اولیه و زیست توده فیتوپلانکتون در مناطق مختلف یک صخره بسیار متفاوت است. در بیشتر موارد، کمترین مقادیر تولید زیست توده و فیتوپلانکتون مربوط به نواحی کم عمق صخره و مخصوصاً مسطح صخره است، جایی که آبی که در طول جزر و مد از روی صخره عبور می کند، بیشترین تماس را با جمعیت کف صخره دارد. صخره دلیل کاهش فیتوپلانکتون در نواحی کم عمق تخت را باید در مصرف آن توسط فیدرهای فیلتر پایین صخره دید (Glynn, 1973a). علاوه بر این، سلولهای dnatoms، مانند Nitzschia delicatissima یا سلولهای شاخهدار Chaetoceros، میتوانند در آبهای متلاطم صخره آسیب ببینند، بهویژه هنگام برخورد با سطح سخت صخره و با ذرات معلق. در آبهای قسمتهای عمیقتر تالاب یا فراتر از لبه بیرونی صخره، جایی که تماس آب با صخره کاهش مییابد، رشد فیتوپلانکتونها به دلیل انتشار برخی مواد مغذی در جوامع پایینی صخره تشدید میشود.
از نظر شاخص های کمی زیست توده و تولید آن، و از نظر غلظت کلروفیل، فیتوپلانکتون در منطقه صخره غنی تر از آب های اطراف اقیانوس باز است (Michel, 1969؛ Sorokin, 1971؛ Ricard, 1976؛ Sournia, Ricard. ، 1976a). شکل 6.3 یک تصویر معمولی از تغییرات در تولید اولیه فیتوپلانکتون در امتداد بخش هایی از صخره هرون (B. Barrier Reef) تا اقیانوس باز ارائه می کند. قبلاً در فاصله 6-9 کیلومتری بیش از یک مرتبه قدر کاهش می یابد.
شفافیت آب در امتداد دیسک سفید در نواحی عمیق صخره ها معمولاً بسیار زیاد است و به طور متوسط 10-15 متر است، اگرچه در برخی موارد می تواند کمتر (5 متر در آتول کانتون) و بیش از 30 متر باشد (ماجورو آتول). ). بنابراین، فتوسنتز فیتوپلانکتون ها عملاً توسط نور در کل ضخامت آنها محدود نمی شود. در شکل شکل 6.11 منحنی های میرایی معمولی را برای تشعشعات فرودی در آب های برخی از صخره های اقیانوس هند با شفافیت متوسط نشان می دهد (10-18). همانطور که از این شکل مشاهده می شود، فیتوپلانکتون ها می توانند کمبود نور را فقط در انتهای تالاب تجربه کنند.
نمونه هایی از تنوع فصلی فیتوپلانکتون ها در شکل 1 ارائه شده است. 6.9،6.12،6.13. آنها نشان می دهند که در فصول مختلف سال نه تنها ترکیب آن تغییر می کند (نگاه کنید به 6.2.1)، بلکه تغییراتی در زیست توده و فعالیت عملکردی آن وجود دارد. نوسانات در مقادیر تولید فیتوپلانکتون اولیه و غلظت کلروفیل در صخره ها در طول فصل به مرتبه بزرگی رسید (Angot، 1968؛ Sournia، 1972). علاوه بر این، حتی در تالاب آتول تاکاپوتو، واقع در منطقه ای از آب های الیگوتروف، غلظت کلروفیل در طول حداکثر فصلی به سطح مشخصه آب های مزوتروف رسید (سورنیا و ریکارد، 1976). در آبهای سد و صخرههای ساحلی، غلظت آن نزدیک به 1 میلیگرم بر متر مکعب بود که با محتوای آن در آبهای اوتروفیک مطابقت دارد و زیست توده فیتوپلانکتون در منطقه صخرههای جزیره نوسی به بیش از 3 گرم در متر مکعب بود. نوسانات فصلی قابل توجهی در غلظت کلروفیل در آب (از 0.13 تا 0.5-2.5 mg/m3) در تالاب B. از صخره سد استرالیا در نزدیکی تاونزویل (Sammarco, Crenshaw, 1984) مشاهده شد. حداکثر در ماه های تابستان-پاییز از دسامبر تا آوریل مشاهده شد. با یک دوره باران و افزایش سطح زهکشی مصادف شد (شکل 6.14). در این دوره، رشد شدید فیتوپلانکتون ها در آب های تالاب مشاهده شد که بخش قابل توجهی از آن جلبک سبز آبی Trichodcsmium بود (Relevante et al., 1982). اثر محرک جریان رودخانه بر توسعه فیتوپلانکتونها برای آبهای صخرهای خارج از کالدونیای جدید نیز مورد توجه قرار گرفت (بینه، 1984). داده های ارائه شده بیانگر بی اساس بودن این نظریه است (پارسونز و همکاران، 1977) که جوامع پلانکتون در آب های گرمسیری، بر خلاف آب های معتدل، در حالت نوعی هموستاز هستند و تغییرات فصلی قابل توجهی را تجربه نمی کنند. چنین ایده هایی ظاهراً نتیجه فقدان چرخه های مشاهده فصلی مناسب در آب های گرمسیری بود (سورنیا، 1969). دلیل ماکزیمم های فصلی در توسعه فیتوپلانکتون ها در آب های استوایی ساحلی در درجه اول به دلیل تغییرات در گردش آب در منطقه اقیانوسی مجاور صخره ها است که دوره تناوب سالانه دارند. علاوه بر این، این تغییرات، حتی در مناطق گرمسیری، اغلب در پاییز و بهار رخ می دهد. تصادفی نیست که حداکثرهای فصلی رشد فیتوپلانکتون ها در پاییز یا بهار مشاهده می شود.
در جریان مطالعه توسعه فیتوپلانکتون در آبهای صخره، وجود نوسانات روزانه قابل توجه در ترکیب و فراوانی جوامع آن و همچنین بهره وری آن مشخص شد (میلیمن و مهنکن، 1972؛ سورنیا و ریکارد، 1976؛ سوروکین). ، 1979). چنین نوساناتی در جوامع فیتوپلانکتون اقیانوس باز نیز رخ می دهد. آنها در آنجا به دلیل تناوب روزانه رشد فیتوپلانکتون ها و تشدید مصرف جلبک ها در شب توسط زئوپلانکتون ها که از لایه های زیرین آب به منطقه یوفوتیک مهاجرت می کنند، ایجاد می شوند. همین دلایل در آبهای ساحلی در منطقه صخرههای مرجانی عمل میکنند و باعث تناوب روزانه در توسعه فیتوپلانکتونها میشوند. اما در این آب ها تحت تأثیر چرخه جزر و مد روزانه نیز قرار دارند. همانطور که نتایج مشاهدات روزانه بر روی صخرههای نزدیک جزایر نامزو نشان داده است، تحت تأثیر چرای فیتوشانکتونهایی که شبها توسط زئوپلانکتونهای صخرهای به ستون آب مهاجرت میکنند، تولید روزانه آن در اوایل صبح با یک مرتبه قدر کاهش مییابد. . با این حال، تا ساعت 11 صبح، جمعیت فیتوپلانکتون تا حد زیادی بازسازی می شود، به خصوص در لبه صخره، جایی که فیتوپلانکتون به شدت تولید مثل می کند (شکل 6.15).
دلایل افزایش بیومس و فیتوتانکتون در آبهای صخره ای در مقایسه با آبهای اطراف اقیانوس آزاد چیست؟ این سوال کاملاً پیچیده است، به ویژه با توجه به این واقعیت که چنین افزایشی در غلظتهای مطلق بسیار کم مواد مغذی رخ میدهد، که تفاوت چندانی با موارد موجود در اقیانوس باز ندارد، و همچنین در شرایط بهرهبرداری غیرمعمول فشرده از جوامع فیتوپلانکتون توسط فیلتر. تغذیه جانوران صخره. نظرات بیان شده در این مورد بسیار متناقض است (Sournia, Ricard, 1976; Sournia, 1976b; Salvat 1982). اکثر نویسندگان به اصطلاح "اثر توده ای جزیره" به عنوان دلیل اصلی اشاره می کنند. این اثر شامل غنیسازی آبهای منطقه ساحلی جزایر با مواد مغذی به دلیل تعامل جرم آنها با جریان ورودی جریانهای اقیانوسی و در نتیجه بالا آمدن آبهای عمیق به سطح است. تأثیر غنیسازی ناحیه نریتی بر جزایر واقع در هسته جریانهای باد تجاری، مانند جزایر مارکیز (Sournia, 1976c). B. دیواره مرجانی (Gilmartin، Relevante، 1974)، هاوایی (Doty، Capurro، 1959)، سیشل (مشاهدات ما) بدون شک رخ می دهد. تحت تأثیر آن، تولید اولیه در منطقه نریتیک چنین جزایری بسیار زیاد است، که بر سطح بهره وری خود آبهای صخره تأثیر می گذارد. با این حال، حتی در این شرایط، تولید و زیست توده فیتوپلانکتون در آبهای تالابهای صخرهای معمولاً بیشتر از آبهای منطقه نریتیک مجاور آنها و در حالت تبادل شدید با آنها است (مارشال، 1965؛ میشل، 1969). ؛ سورنیا، ریکارد، 1976، 1976 الف؛ تامپسون، گلدینگ، 1982). با توجه به این داده ها، می توان نتیجه گرفت که دلیل اصلی افزایش بهره وری فیتوپلانکتون در آب های صخره ای، انتشار مداوم عناصر بیوژنیک (عمدتاً نیتروژن، فسفر) به ستون آب توسط هتروتروف های کف و پلانکتون است که به عنوان یک نتیجه فرآیندهای تخریب مواد آلی است که در صخره ها با نیروی زیاد رخ می دهد. در عین حال، به دلیل سرعت بسیار بالای گردش مواد مغذی در ستون آب صخرهها، همیشه نمیتوان افزایش غلظت مطلق مواد مغذی نمک را در آبهای منطقه ریف نسبت به اقیانوس مجاور ثبت کرد. آب (شکل 5.1.).